Tutte le attività
aggiornamento automatico
- Ultima settimana
-
 FABRIZIO1967 si è unita/o alla comunità
FABRIZIO1967 si è unita/o alla comunità -

Platanthera chlorantha (Custer) Reichenbach 1828
bgiordy ha postato un topic nell'area Orchidee Veneto Anno 2026
Platanthera chlorantha (Custer) Reichenbach 1828 - Colli Berici - 247 m - 18 mag 2026
-
Anacamptis palustris (L.) R.M. Bateman, Pridgeon & M.W. Chase 1997
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026

-
Anacamptis palustris (L.) R.M. Bateman, Pridgeon & M.W. Chase 1997
bgiordy ha postato un topic nell'area Orchidee Veneto Anno 2026
Non c'erano più di una decina di piante. E pensare che una volta il prato era pieno. Stanno facendo la stessa fine di A laxiflora. Anacamptis palustris (L.) R.M. Bateman, Pridgeon & M.W. Chase 1997 - Campagna Lupia - 1 m - 17 mag 2026
-
Anacamptis ×duquesnei (Rchb.f.) H.Kretzschmar, Eccarius & H.Dietr. 2007
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026

-
Anacamptis ×duquesnei (Rchb.f.) H.Kretzschmar, Eccarius & H.Dietr. 2007
bgiordy ha postato un topic nell'area Orchidee Veneto Anno 2026
Anche quest'anno c'erano due ibridi. Anacamptis ×duquesnei (Rchb.f.) H.Kretzschmar, Eccarius & H.Dietr. 2007 - (A. palustris × A. pyramidalis) - Campagna Lupia - 1 m - 17 mag 2026
-

Ophrys apifera Huds. 1762.
Giuliano Gnata ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
Grazie Giordano, ma quando si deve andare si và. - Precedente/i
-
Ophrys apifera Huds. 1762.
bgiordy ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
Il fango ti sarà arrivato alle orecchie, con tutte queste piogge ! Certo che sei coraggioso ad andare in giro con questo freddo ! Valeva la pena per queste bellezze!! -
Dopo le forti piogge un giro al M. Summano è di dovere. Ophrys apifera Huds. 1762.





-
Ophys apifera var. trollii micro-labellata
Giuliano Gnata ha risposto alla discussione di fiorenzolovison in Orchidee Veneto Anno 2026
Spettacolo! -

Pholiota adiposa (Batsch : Fr) P. Kumm. 1871
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
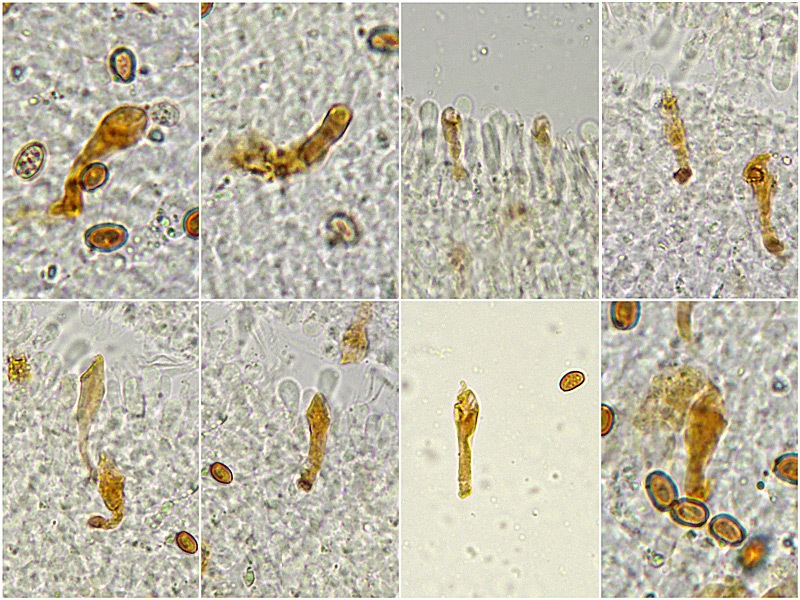
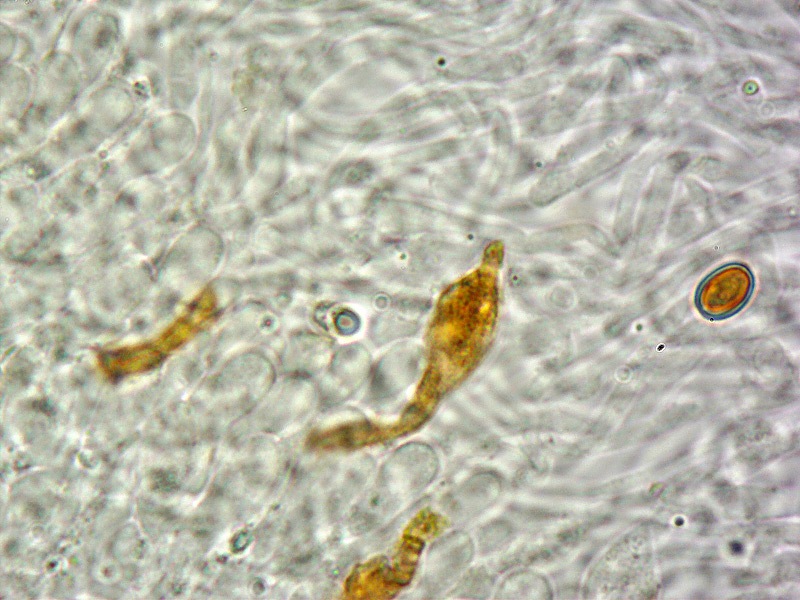
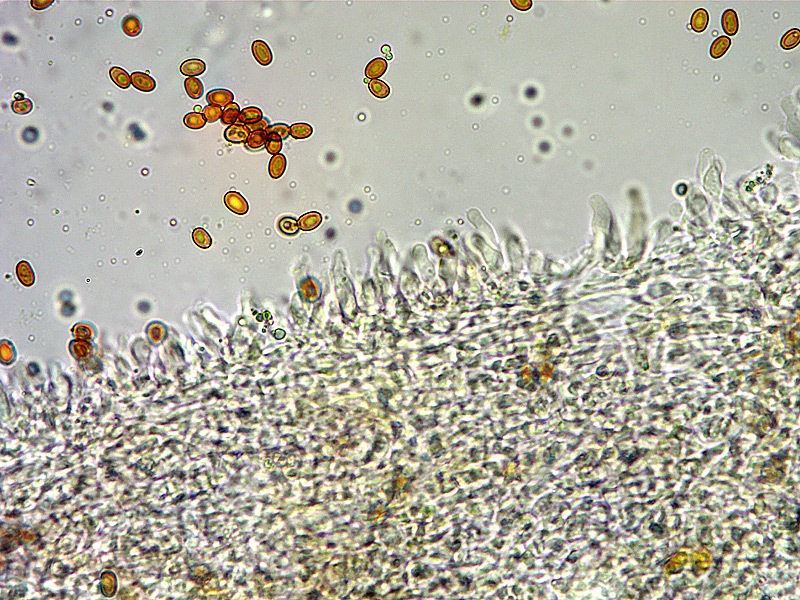
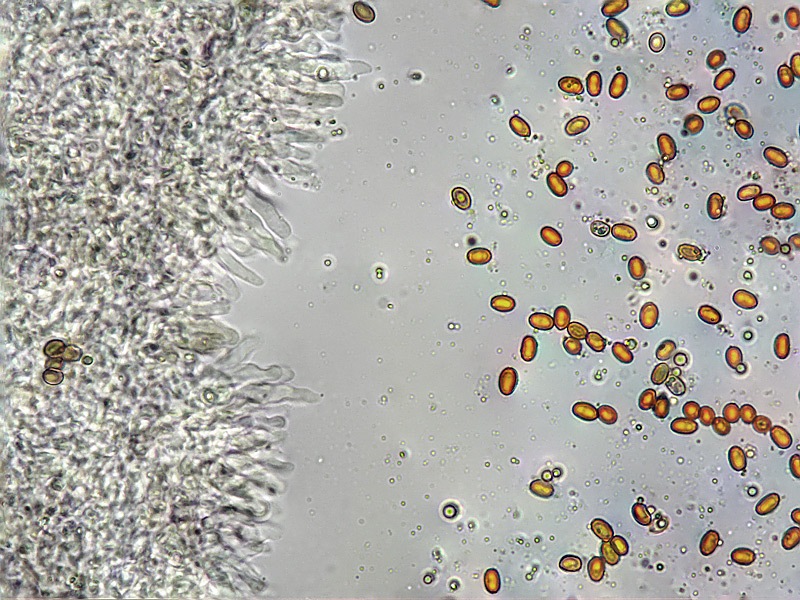
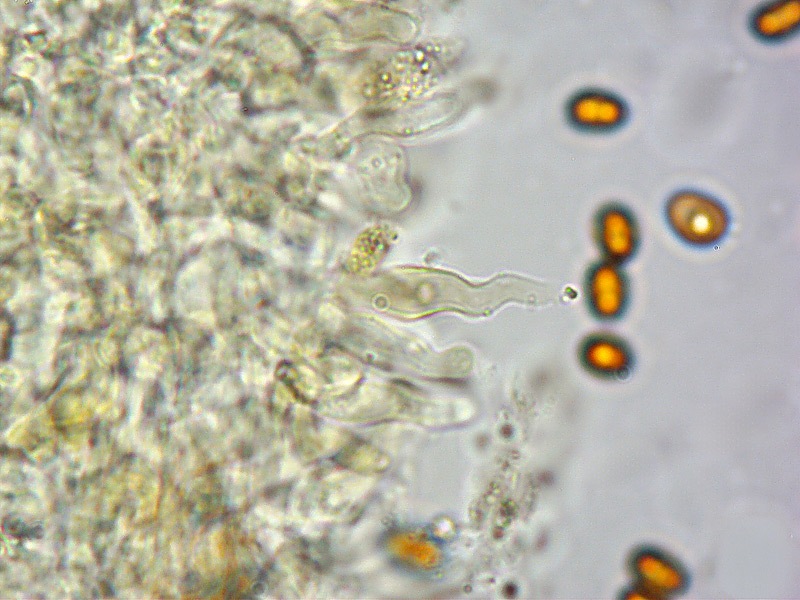
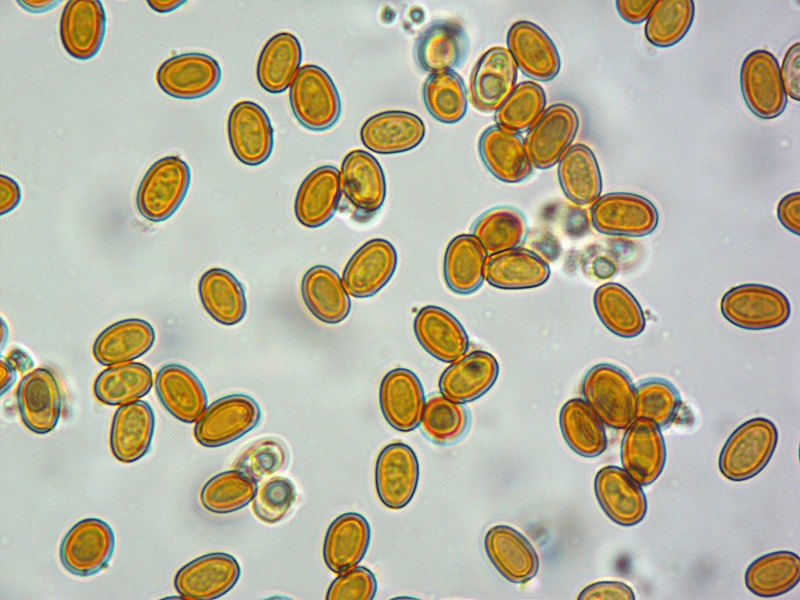
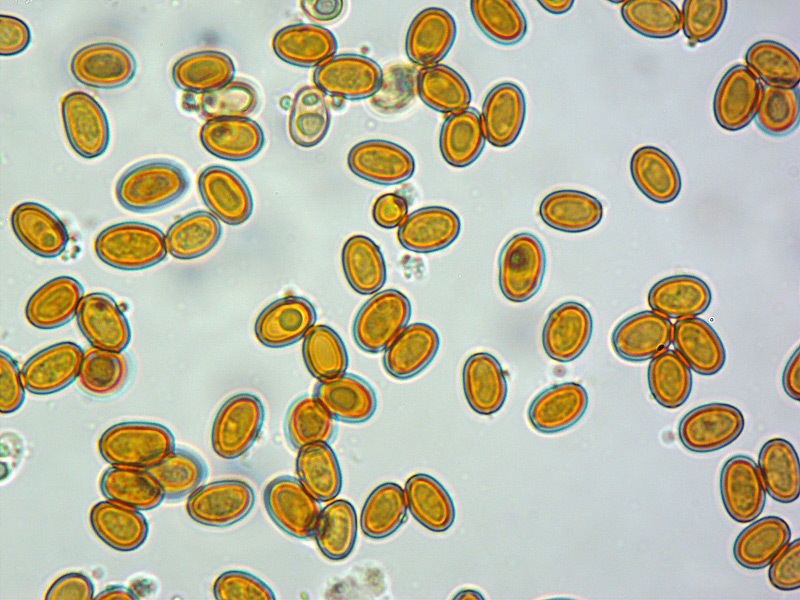
Pholiota adiposa (Batsch : Fr) P. Kumm. 1871 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Sinonimi Pholiota lilacifolia P.D. Orton 1976 Pholiota aurivella var. abietis-nordmanniana Singer 1930 Note tassonomiche Alcuni autori (AA.VV. 2012) ritengono questa specie sinonimo prioritario di Pholita aurivella (Batsch : Fr.) P. Kumm. s. auct p.p., sinonimia non condivisa da tutti gli autori. Etimologia L'epiteto Pholiota dal greco φολίς [pholís] = squama e da οὖς [otós] = orecchio, per la presenza di squame sul carpoforo. L'epiteto adiposa deriva dal latino ădeps = grasso, per la superficie del cappello viscida. Cappello 20-150 mm, emisferico, più disteso a maturità, con il margine involuto, presenta una cuticola, viscida, appiccicosa, di color giallo oro, poi bruno aranciato, con presenza di squame di colore marrone rossastro, non particolarmente rialzate, non echinulate. Imenoforo Lamelle mediamente fitte, da adnate a subdecorrenti, alternate a lemellule di varie dimensioni, inizialmente di colore crema ocracee, il filo lamellare è finemente fimbriato e sterile per la presenza di cheilocistidi, discolore rispetto alle facce lamellari. Anello Evidente, bianco, fibroso, membranaceo, ben evidente già nei soggetti molto giovani, spesso lembi abbondanti del velo secondario rimangono attaccati al bordo del cappello. Gambo Cilindrico, appuntito e spesso curvo alla base, squamoso su tutta la superficie inferiore all'anello. Carne Giallastra, odore insignificante, fungino; sapore neutro. Habitat Cespitoso, fruttifica su legno soprattutto di Fagus sylvatica, raramente su legno di conifere. Microscopia Cheilocistidi di profilo vario, ma non capitulati: strettamente clavati, lageniformi, serpeggianti. Pleurocrisocistidi spesso con forme irregolari, a volte muniti di una protuberanza apicale (mucronati), distribuiti nella trama lamellare. Basidi tetrasporici, clavati. Spore (6,9) 7,9-8,7 (9,7) × (4,8) 5,1-6,0 (6,6) µm; Q = (1,3) 1,4-1,6 (1,7); N = 50; Media = 8,3 × 5,5 µm; Qm = 1,5; ellissoidali, lisce a parete spessa, provviste di un evidente poro germinativo. GAF abbondanti, presenti in tutti i tessuti. Commestibilità e tossicità Non commestibile. Specie simili Pholiotha adiposa si caratterizza per la presenza di squame marroni sul cappello, presenza di cheilocistidi, pleurocrisocistidi, e per le spore con misure medie maggiori di 7 × 5 µm. Pholiota cerifera (P. Karst) P. Karst è la specie più simile a Pholiota adiposa, si distingue per il cappello non fortemente glutinoso, i chilocistidi clavati, ma più larghi di quelli di Pholiota adiposa e i crisocistidi abbondanti. Pholiota limonella (Peck) Sacc. è difficilmente distinguibile dal punto di vista dei caratteri macroscopici da Pholiota adiposa. Cresce su conifere e latifoglie ma difficilmente su Fagus sylvatica, ha spore più strette (4,0-5,2 (5,5) µm) di Pholiota adiposa ((4,5) 5,0-6,2 (6,5) µm). Pholiota jahnii Tjallingii-Beukers & Bas si distingue per le squame più appuntite e nerastre sul cappello, per le spore ellissoidali, ovoidali, più piccole (5-7 × 3-4 µm). Pholiota squarrosoides (Peck) Sacc. si distingue per le spore ellissoidali, faseoliformi in vista laterale, più piccole (4,5-6,5 × 2,5-3,5 µm). Pholiota lundbergii Jacobsson ha crescita terricola e spore più piccole (5,5-7,5 × 3,5-4,3 µm). Bibliografia AA.VV., 2012. Funga Nordica. Agaricoid, boletoid and cyphelloid genera. Ed. Nordsvamp. HOLEC, J., 2001. The Genus Pholiota in central and western Europe. Libri Botanici N° 2. Scheda di proprietà AMINT realizzata da Tomaso Lezzi - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Lazio. Maggio 2026. Foto di Tomaso Lezzi. Crisocistidi. Osservazione in KOH, a 600×. Crisocistidio. Osservazione in KOH, a 1000×. Cheilocistidi. Osservazione in KOH, a 400×. Crisocistidi. Osservazione in KOH, a 1000×. Spore. Osservazione in KOH, a 1000×.-03.jpg.1354008db92101376ecb4cffa4be42e5.jpg)
-04.jpg.e76d9665cc708b8a6b7a8254f1a9d8ef.jpg)
-05.jpg.18654a2e5855af15482170b369a66507.jpg)
-
Ophrys apifera var. trollii (Hegetschw.) Rchb. 1851
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Ho notato anch'io che in Friuli sono parecchio diverse: semplice variabilità? Boh! -
Ophys apifera var. trollii micro-labellata
bgiordy ha risposto alla discussione di fiorenzolovison in Orchidee Veneto Anno 2026
Il labello potrebbe anche essere stato mangiato, ma i petali indicano un lusus. Bella colorata di rosa, non l'ho mai incontrata di questo colore. -

Ophys apifera var. trollii micro-labellata
fiorenzolovison ha risposto alla discussione di fiorenzolovison in Orchidee Veneto Anno 2026
seconda piantita
-
Ophys apifera var. trollii micro-labellata
fiorenzolovison ha postato un topic nell'area Orchidee Veneto Anno 2026
trovata in mezzo ad apifere e rare trollii: lusus /regressione?
-
Ophrys apifera var. trollii (Hegetschw.) Rchb. 1851
fiorenzolovison ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
bellissima, ma ogni anno mi domando : è piuttosto diversa dalle Trollii che fotografo in Friuli e quelle presenti in Internet (vedi fot 2026)
-
Ophrys apifera var. aurita (Moggr.) Gremli 1887
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Grazie Giuliano. -
Orchis pallens L.
bgiordy ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
Brava Luisa! -
Himantoglossum adriaticum H. Bauman 1978
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Purtroppo sta succedendo dappertutto. Non bastano i cinghiali . Grazie giuliano. -
Ophrys apifera var. trollii (Hegetschw.) Rchb. 1851
bgiordy ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Grazie Giuliano. -
Ophrys apifera var. trollii (Hegetschw.) Rchb. 1851
Giuliano Gnata ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Deliziose. -
Ophrys apifera var. aurita (Moggr.) Gremli 1887
Giuliano Gnata ha risposto alla discussione di bgiordy in Orchidee Veneto Anno 2026
Bella, dovrei andare a vedere le mie poco lontano da casa. -
Orchis pallens L.
Giuliano Gnata ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
Il merito è di mia moglie Luisa. -
Coeloglossum viride (L.) Hartm. 1820.
Giuliano Gnata ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
-
Orchis militaris L. 1753.
Giuliano Gnata ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
-
Orchis ovalis F.W.Schmidt 1791. in Mayer, Samml.
Giuliano Gnata ha risposto alla discussione di Giuliano Gnata in Orchidee Veneto Anno 2026
Grazie.

