
Archivio Micologico
-
Numero contenuti
15665 -
Iscritto
-
Ultima visita
Tipo di contenuto
Profili
Forum
Orchidee
Diventa Socio
Calendario
Tutti i contenuti di Archivio Micologico
-

Neolentinus schaefferi (Weinm.) Redhead & Ginns 1985
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus schaefferi (Weinm.) Redhead & Ginns; Regione Lombardia; Anno 2005; Foto di Massimo Biraghi.
-
Neolentinus schaefferi (Weinm.) Redhead & Ginns 1985
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Neolentinus schaefferi (Weinm.) Redhead & Ginns 1985 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Polyporaceae Sinonimi Lentinus cyathiformis (Schaeff.) Bres. 1900 Foto e Descrizioni Cresce su legno di caducifoglie, raramente su conifere, lamelle lungamente decorrenti, finissime e tipicamente seghettate. Regione Lombardia; Anno 2005; Foto di Massimo Biraghi.
-
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Lazio, Roma, Villa Ada; Foto di Tomaso Lezzi.


-
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Lazio, Roma, Villa Ada; Foto di Luigi Minciarelli. Crescita singola su tronco marcescente di Pinus pinea L. Dettaglio della lamelle, dal filo tipicamente seghettato. Un particolare del centro della cuticola squamettata. Particolare della base del gambo. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Trentino; Settembre 2011; Foto di Luigi Minciarelli. Un lignicolo dall'odore forte di anice. Imenoforo. Foto di Mario Iannotti. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Trentino; Agosto 2011; Foto di Tomaso Lezzi. 1º Congresso trentino sui funghi alpini, Cles (TN), Loc. Malgaroi. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Trentino; Luglio 2010; Foto di Pietro Curti. Macro del pileo. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Lombardia, Valle Camonica; Maggio 2011; Foto di Lorenzo Martinelli. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Trentino; Luglio 2006; Foto di Mauro Cittadini. Regione Trentino, Settembre 2007 Trentino, foto Piero Curti. -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Lombardia, Spiazzi di Gromo; Luglio 2006; Foto di Massimo Biraghi. (Exsiccatum MB20060710-66) -
Neolentinus lepideus
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fries) Redhead & Ginns; Regione Trentino; Anno 2003; Foto di Pietro Curti. Aggiungo una nuova bella foto di questa specie lignicola, dal forte odore anisato e dal tipico filo lamellare seghettato, semplice la determinazione sul campo.
-
Neolentinus lepideus
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Neolentinus lepideus (Fr.) Redhead & Ginns 1985 Tassonomia Divisione Basidiomycota Ordine Gloeophyllales Famiglia Gloeophyllaceae Genere Neolentinus Sinonimi Lentinus lepideus (Fr.: Fr.) Fr. 1825 Neolentinus lepideus f. ceratoides (Holmsk.) Bondartseva, Zmitr. & Zarudnaya 2016 Note tassonomiche Redhead & Ginns (1985) hanno ricombinato alcune specie di Lentinus nel nuovo genere Neolentinus. La differenza tra i due generi consiste principalmente nella produzione di carie bianca nel substrato legnoso nel genere Lentinus e di carie marrone nel genere Neolentinus. Lo status di genere è stato poi ampiamente accettato a seguito delle ricerche genetiche effettuate da Moncalvo et al. (2002). Etimologia L'epiteto Neolentinus deriva dal prefisso latino neo- = nuovo e Lentinus, nome di genere, poiché il genere Neolentinus è stato separato dal genere Lentinus. L'epiteto lepideus deriva dal greco λεπίς [lepìs] = squama, per il cappello squamoso. Cappello 5-30, da convesso a piano, fino a leggermente depresso o ciatiforme a maturità; superficie da biancastra a sfumata di ocraceo, asciutta, liscia e lucente, tipicamente dissociata in squame scure, distribuite più o meno ad anelli concentrici. Lamelle Smarginate, decorrenti, sinuate, larghe, non anastomosate o venoso congiunte, da bianche a giallo chiaro, con lamellule di varie lunghezze, filo fortemente dentato. Gambo 4–11 x 1–3, centrale o eccentrico, cilindrico con base radicante, di solito inserita in un supporto legnoso. Superficie concolore al cappello, marrone nerastra alla base; con squame concolori o più scure. Presenza di anello nella parte alta, spesso effimero ma a volte persistente. Carne Bianca, sempre più coriacea con l’età; odore molto aromatico, forte, di anice. Habitat Cresce su tronchi tagliati (o radici) di conifere, in particolare di Pinus spp., Abies spp., Picea spp., Larix spp.e Sequoia spp. Occasionali ritrovamenti sono stati effettuati anche su Quercus spp. e Ulmus spp. Microscopia Spore cilindriche, ialine, a parete sottile. Basidi clavati, allungati, bi- e tetrasporici. Cistidi assenti. Filo lamellare sterile con presenza di cellule allungate cistidiformi, sinuose, cilindriche o subclavate, ialine, a parete sottile. Commestibilità o Tossicità Non commestibile, per la consistenza coriacea della carne. Osservazioni Specie distribuita principalmente nelle zone temperate dell’emisfero settentrionale, con un areale che va dall’Europa all’Asia occidentale, fino al Nord America. Il micelio ha un’intensa azione di decomposizione del substrato legnoso, ed è stato più volte responsabile di cadute di pali telegrafici e pali di sostegno nelle miniere. Specie simili Neolentinus lepideus è una specie di facile determinazione grazie al tipico odore di anice (sambuca) e per l’habitat su legno di conifera. Neolentinus ponderosus (O.K. Mill.) Redhead & Ginns la cui distribuzione pare sia limitata alla parte occidentale degli U.S.A. è legato a legno di conifere, in special modo di Pinus ponderosa Dougl. e differisce dal Neolentinus lepideus per la taglia decisamente più grande, fino a 50 cm di diametro, per l'odore non aniseo e per la totale assenza di un velo parziale e quindi di un anello. Bibliografia REDHEAD S.A., GINNS J. 1985. A reappraisal of agaric genera associated with brown rots of wood. Transactions of the Mycological Society of Japan. 26: 349-381 MONCALVO et.al. 2002. One Hundred and Seventeen Clades of Eu-agarics. Molecular Phylogenetics and Evolution 23: 357-400. Scheda di proprietà AMINT realizzata dal CLR Micologico di AMINT. Regione Trentino. Anno 2003. Foto di Pietro Curti.
-
Neobulgaria pura (Pers. : Fr.) Petr. 1921
Archivio Micologico ha postato un topic nell'area Funghi Velenosi
Neobulgaria pura (Pers. : Fr.) Petr. 1921 Tassonomia Divisione Ascomycota Classe Leotiomicetes Ordine Helotiales Famiglia Leotiaceae Genere Neobulgaria Sinonimi Peziza pura Pers. : Fr. 1796 Bulgaria pura (Pers. : Fr.) Fr. 1822 Ombrophila pura (Pers. : Fr.) Baral 1985 Etimologia Dal greco neos, νέος [-α, -ον] = nuovo e dal latino bulga = borsa di cuoio o pelle di otre, per l’aspetto di una borsa di cuoio in riferimento al precedente genere Bulgaria, istituito da Fries prendendo spunto dal preesistente genere Burcardia. Dal latino purus, ă, um = puro, trasparente. Ascoma Apotecio di forma per lo più turbinata con diametro di circa 2-5 mm che tende a svilupparsi in piccoli agglomerati di ascomi appressati fra loro che gli conferiscono, nel complesso, un aspetto tremelloide. Presenta consistenza decisamente gelatinosa e lievemente appiccicosa, aspetto traslucido e tonalità rosa-isabella in tutto l'apotecio. L'imenoforo è liscio e concavo mentre la superficie esterna è rugosa e lievemente granulosa. Carne Biancastra, inodore. Habitat Cresce in gruppi di numerosi individui prevalentemente su legno marcescente di Fagus, talvolta di Alnus. Microscopia Aschi 88-92 × 6,9-7,8 µm con estremità amiloide, octosporici, cilindrico-clavati. Spore 6,2-7,7 × 3,2-3,8 µm; Q = 1,7-2,4; Qm = 2,0 ellissoidali, lisce, ialine, biguttulate, solitamente uniseriate nell'asco. Parafisi cilindracee, settate e sottili. Commestibilità e tossicità Sindrome Giromitrica, contenuto irrilevante di Giromitrine tali da non costituire un pericolo reale. Non esistono casi clinici segnalati. Potenziale pericolo se consumate crude o poco cotte in grandi quantità. Somiglianze e varietà Ascotremella faginea (Peck) Seaver si presenta con apoteci globosi che rendono un aspetto complessivo cerebriforme. Ha spore molto simili in forma e dimensioni, ma leggermente striate in senso longitudinale. Ascocoryne sarcoides (Jacq. : Fr.) J. W. Groves & D.E. Wilson è di color viola scuro e presenta spore più grandi (15 × 4,5 µm). Ascocoryne cylichnium (Tul.) Korf è di color porpora e ha spore decisamente più grandi (30 × 6 µm). Ascocoryne turficola (Boud.) Korf è di color rosa vinoso, presenta uno stipite più o meno allungato, ha crescita sfagnicola e possiede spore di dimensioni maggiori (14-19,6 × 5,6-7 µm). Neobulgaria pura var. foliacea (Bres.) Dennis & Gamundí ha spore leggermente più grandi (6,5-9 × 3,5-4,5 µm) e aspetto globoso-cerebriforme. Bibliografia LIZON, P., ITURRIAGA, T., KORF, R.P., 1998. A preliminary discomycete flora of Macaronesia: Part 18, Leotiales. Mycotaxon LXVII: 73-83. MEDARDI, G., 2006. Atlante fotografico degli Ascomiceti d'Italia. Ed. AMB. MOORE, E.J., 1965. Ontogeny of Gelatinous Fungi. Mycologia LVII, 1: 114-130. STASIÑSKA, M., SOTEKA, Z.,2004. Ascocoryne turficola (Fungi, Ascomycetes), a species new to Poland. Acta Societatis Botanicorum Poloniae LXXIII, 1: 61-64. Scheda di proprietà AMINT realizzata da Francesco Tozzi - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Abruzzo, Bosco Scaricaturo, Palena (AQ); Ottobre 2018; Foto, descrizioni e microscopia di Francesco Tozzi. Esemplari rinvenuti su legno marcescente di Fagus sylvatica. Aschi 88-92 × 6,9-7,8 µm con estremità amiloide, octosporici, cilindrico-clavati. Parafisi sottili, cilindracee, leggermente più lunghe degli aschi. Spore 6,2-7,7 × 3,2-3,8 µm ellissoidali, biguttulate. Osservazione in Melzer a 600×. Parafisi e aschi con spore monoseriate e biguttulate. Osservazione in Melzer a 1000×.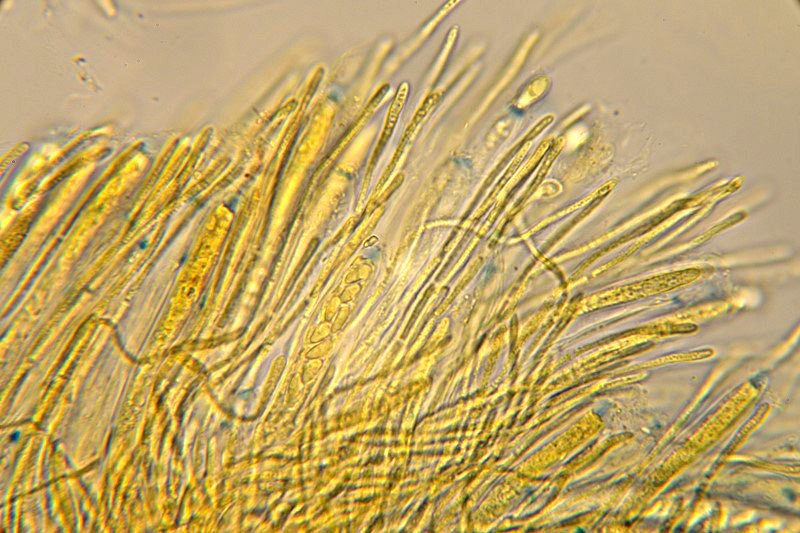
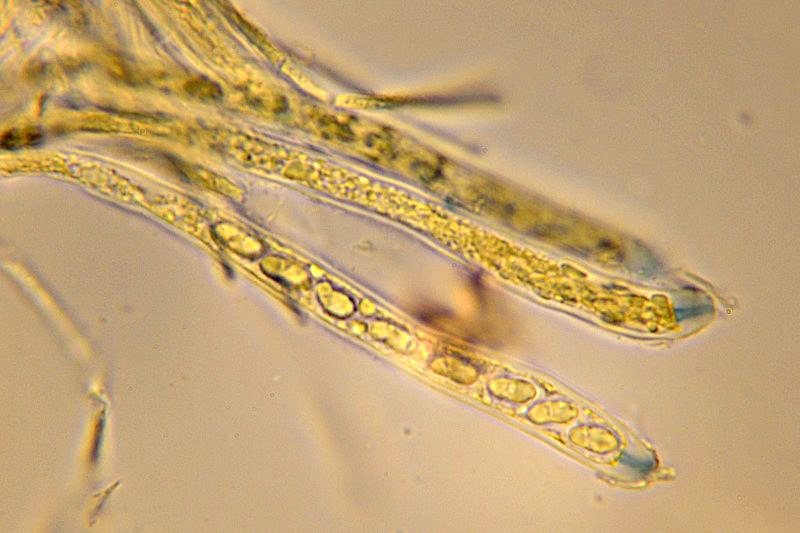
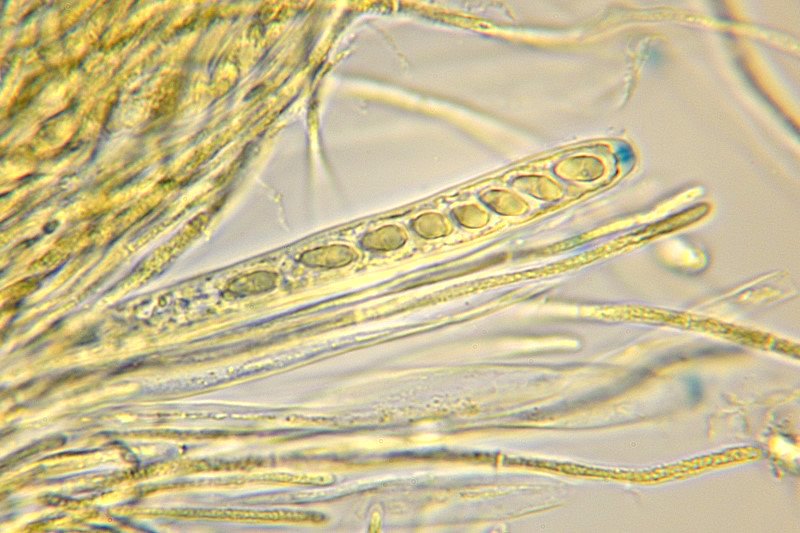
-
Nectria peziza (Tode : Fr.) Fr. 1849
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Nectria peziza (Tode : Fr.) Fr.; Regione Lazio; Gennaio 2010; Foto di Felice Di Palma. -
Nectria peziza (Tode : Fr.) Fr. 1849
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Nectria peziza (Tode : Fr.) Fr. 1849 Tassonomia Divisione Ascomycota Classe Sordariomycetes Ordine Hypocreales Famiglia Nectriaceae Nome italiano Non risultano nomi volgari per questa specie. Etimologia Néctria: dal greco nektrìs, nuotatrice, per l'habitat su legno bagnato, intriso di acqua. peziza: per somiglianza, a fine ciclo, alla forma di tazza o catino tipica degli ascomi del genere Peziza. Sinonimi Cucurbitaria peziza (Tode) Kuntze 1898 Dialonectria aurea (Grev.) Cooke 1884 Neuronectria peziza (Tode) Munk 1957 Ascoma Globoso, finemente asperulato, color rosso-arancio intenso, in media con diametro di 0,4 mm; crescita in esemplari singoli, non raggruppati in stroma. In fase di maturazione i periteci formano in zona apicale una papilla. Successivamente la papilla si fora a formare un ostiolo dal quale fuoriescono le spore addensate in filamenti biancastri. Al terime del ciclo di dispersione delle spore, il peritecio si sgonfia, collassa assumendo la tipica forma a coppetta che ha richiamato per similitudine il nome specifico di peziza. Carne Sottilissima, di consistenza prima abbastanza dura, poi elastica e infine fragile alla rottura. Microscopia Spore (11)11,5-14(15) × 5,5-6,5 µm, ialine al microscopio ottico, lisce se osservate in acqua o Rosso Congo acquoso; quelle mature appaiono nettamente striate o comunque finissimamente sabbiate se colorate in Blu Cotone Lattico, settate in modo netto con lieve strozzatura mediana, guttula/e presente/i in ciascuna delle due parti. Aschi 82-102 × 9,5-12 µm, ottosporici, con parete sottile, parafisi non rilevate. Trama del peritecio con struttura globoso-angularis. Habitat Su legno decorticato, degradato, molto umido, imbibito d'acqua, di latifoglia nel caso del nostro ritrovamento al quale si riferiscono i dati macro e micro riportati. In letteratura viene spesso segnalata su basidiomi di vecchie Polyporales s.l. marcescenti. Osservazioni Da un punto di vista macroscopico, la determinazione può essere agevolata osservando la classica forma 'pezizoide' dei periteci che assumono al termine della fase di espulsione delle spore e la crescita in individui singoli e non raggruppati in stroma. Nella microscopia, il carattere maggiormente distintivo è quello delle striature longitudinali presenti nelle spore mature, rilevabili con l'osservazione in Blu Cotone Lattico. Molte Nectria sp. costituiscono dei pericolosi patogeni per le piante superiori. Di questi funghi viene generalmente distinto lo stadio teleomorfo (stato sessuale, perfetto) da quello anamorfo (forma imperfetta, asessuata) come ad esempio in Nectria cinnabarina (Tode ex Fr.) Fr., stadio teleomorfo che ha come teleomorfo Tubercularia vulgaris Tode. Commestibilità e tossicità Senza nessun interesse alimentare. Specie simili Molte delle Nectria sp. condividono caratteri comuni come habitat, forma e colorazioni rosso aranciate. Dalla comune Nectria cinnabarina (Tode ex Fr.) Fr. si distingue per la crescita isolata e non in densi gruppetti riuniti in stroma e per i caratteri microscopici. Bibliografia AA.VV. (2000). Nordic Macromycetes, Volume 1. Ascomycetes. Ed. L. Hanse and H. Knudsen. Medardi, G. (2005). Atlante fotografico degli Ascomiceti d'Italia. Ed. A.M.B. Cetto, B. (1993). I funghi dal vero, Vol. 7°. Ed. Saturnia. Trento. Lanconelli, L., Ballanti, E., Rava, M. (1998). Funghi del Lughese. Oltre 360 specie di campagna e di città. Ed. Faenza Scheda di proprietà AMINT realizzata da Mauro Cittadini - Approvata e Revisionata dal CLR Micologico di AMINT Nectria Peziza (Tode) Fr.; Foto Mauro Cittadini. Aschi con ascospore settate osservate in rosso Congo, foto Mauro Cittadini. Spore in blu Cotone Lattico, si notano le sottili striature longitudinali, foto Mauro Cittadini. Struttura del peritecio, foto Mauro Cittadini. Particolari ingranditi degli ascomi, si nota la fuoriuscita delle spore dall'ostiolo apicale e qualche peritecio collassato, foto Mauro Cittadini. -
Nectria cinnabarina (Tode) Fr. 1849
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Nectria cinnabarina (Tode) Fr.; Regione Umbria; Novembre 2014; Foto e microscopia di Stefano Rocchi. Aschi. Osservazione in Mercurocromo 400×. Osservazione in Mercurocromo 1000×. Spore ellissoidali-fusiformi, monosettate, lisce e con strozzatura centrale 11,2-18,9 × 4,3-6,7 µm; Qm = 2,5. Osservazione in Mercurocromo 1000×. La struttura del peritecio. Osservazione in Mercurocromo 400×. -
Nectria cinnabarina (Tode) Fr. 1849
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Nectria cinnabarina (Tode) Fr. 1849 Tassonomia Divisione Ascomycota Classe Sordariomycetes Ordine Hypocreales Famiglia Nectriaceae Descrizione Su canna di bambù marcescente, il fungo è costituito da piccolissimi carpofori rosso acceso di dimensioni inferiori al millimetro e raggruppati tra di loro. Le spore sono chiare, ellissoidali-fusiformi (con leggera strozzatura centrale, tanto da sembrare un'orma di scarpa) lisce con interno oleoso e monosettate 12,2-18,2 × 5-7,7 µm; Qm = 2,4. Aschi ottasporici 76,9-85,1 × 7-7,8 µm. Parafisi assenti. Altre Nectria di colore rosso vivo sono N. coccinea la quale ha spore più piccole e leggermente verrucose e N. fuckeliana sempre con spore più piccole, la quale si associa di solito a rami di conifere. Regione Umbria; Novembre 2014; Foto e microscopia di Stefano Rocchi. Aschi ottasporici 76,9-83,1 × 7-7,8 µm. Parafisi assenti. Osservazione in Mercurocromo 400×. Osservazione in Mercurocromo 1000×. Spore chiare, ellissoidali-fusiformi (con leggera strozzatura centrale) lisce, con interno oleoso e monosettate 12,2-18,2 × 5-7,7 µm; Qm = 2,4. Osservazione in Mercurocromo 1000×. -
Naucoria zonata E. Ludw. & Reil 2001
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Naucoria zonata E. Ludw. & Reil 2001 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Foto e descrizione Cappello di piccole dimensioni, massimo 2 cm, colorazioni bruno rossastre, più marcate al centro, dove è presente un piccolo umbone, decisamente schiarente all'ocra chiaro a partire da metà raggio che gli dona un aspetto zonato, poco o per nulla igrofano e ricoperto da resti di velo. Lamelle ocracee, scurenti con l'età, gambo cilindrico, un poco bulbosetto, da color miele a subconcolore al pileo e ricoperto da fibrille più chiare più evidenti al terzo superiore. Odore leggermente rafanoide, pungente, con nota di foglie di geranio stropicciate, sapore nettamente amaro. Osservazioni Nelle specie con cheilocistidi fusiformi e/o a pelo d'ortica questa specie poco conosciuta si riconosce per essere l'unica Naucoria con spore di dimensioni minute rispetto alle sue simili. Microscopia Basidi tetrasporici. Cheilocistidi urticoidi (a pelo di ortica) con giunti a fibbia. Spore amigdaliformi - subfusiformi, rugose, con verruche piatte anche marcate. 60 osservazioni N° Long. Larg. Q Moy 6,86 3,99 1,72 Min 6,30 3,44 1,59 Max 7,40 4,30 1,97 Media 6,81 4,07 1,69 Long.: N = 30; dMd; (6,14)6,418-6,95-7,457(7,82) Larg.: N = 30; dMd; (3,55)3,773-3,98333333333333-4,447(4,60) Q: N = 30; dMd; (1,51)1,564-1,64-1,878(1,90) N° Long. Larg. Q Moy 7,00 4,10 1,71 Min 6,14 3,55 1,51 Max 7,82 4,60 1,90 Media 7,06 4,15 1,70 Lxl: N = 30; dMd; (6,14)6,42-6,95-7,46(7,82) × (3,55)3,77-3,98333333333333-4,45(4,60); Qm= 1,71 Regione Lombardia; Novembre 2013; Foto, descrizione e microscopia di Massimo Biraghi. Pianura bergamasca, località Pumengo. Essenze presenti Farnia, Frassino, Nocciolo, nel muschio. Microscopia Basidi tetrasporici. Cheilocistidi urticoidi (a pelo di ortica) con giunti a fibbia. Spore amigdaliformi - subfusiformi, rugose, con verruche piatte anche marcate. 60 osservazioni. N° Long. Larg. Q Moy 6,86 3,99 1,72 Min 6,30 3,44 1,59 Max 7,40 4,30 1,97 Media 6,81 4,07 1,69 Long.: N = 30; dMd; (6,14)6,418-6,95-7,457(7,82) Larg.: N = 30; dMd; (3,55)3,773-3,98333333333333-4,447(4,60) Q: N = 30; dMd; (1,51)1,564-1,64-1,878(1,90) N° Long. Larg. Q Moy 7,00 4,10 1,71 Min 6,14 3,55 1,51 Max 7,82 4,60 1,90 Media 7,06 4,15 1,70 Lxl: N = 30; dMd; (6,14)6,42-6,95-7,46(7,82) × (3,55)3,77-3,98333333333333-4,45(4,60); Qm= 1,71 -
Naucoria subconspersa Kühner ex P.D. Orton 1960
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Naucoria subconspersa Kühner ex P.D. Orton 1960 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Sinonimi Alnicola subconspersa (Kühner ex P.D. Orton) Bon 1979 Cappello cannella, terra d'ombra, a volte un poco rossastro, igrofano (alutaceo con tempo secco) minutamente squamuloso furfuraceo e ricoperto da un velo nei giovani esemplari; lamelle cannella, larghe, smarginate; gambo pallido in alto, bruno bistro altrove e minutamente flocculoso. Spore 10-13 × 5-6 µm, ovato-ellissoidi, punteggiate. Cistidi a pelo d'ortica con prolungamento apicale più o meno corto e con base rigonfia. Simile a Naucoria scolecina (Fr.) Quél. si distingue per il netto velo e il cappello squamuloso. Regione Lombardia, Gera d'Adda; Novembre 2016. Foto, descrizione e microscopia di Massimo Biraghi. Ritrovamento ai bordi di una roggia con essenze diverse, Ontano, Quercia, Acero campestris, Carpino. Spore ovoidi-ellissoidali, anche subamigdaliformi, verrucose. Osservazione a 400×. N° Long. Larg. Q Moy 9,74 5,44 1,80 Min 8,60 4,97 1,52 Max 10,95 6,24 2,01 Media 9,70 5,41 1,80 Osservazione a 1000×. Basidi tetrasporici. Cheilocistidi. Osservazione a 400×. Moy 38,13 11,38 Min 31,74 6,99 Max 44,85 15,39 Media 37,15 11,65 Osservazione a 1000×. -
Naucoria striatula P.D. Orton 1960
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Naucoria striatula P.D. Orton 1960 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Sinonimi Alnicola striatula (P.D. Orton) Romagn. 1979 Foto e Descrizioni Ritrovamento in bosco planiziale percorso da risorgive, essenze primarie Quercia, Carpino, Ontano, quattro stazioni sulle sponde o nelle immediate vicinanze delle rogge, fruttificazione gregaria, anche abbondate in numerosi esemplari, terreno ricco di sostanze organiche, fogliame, piccoli detriti legnosi, terreno tipico delle sponde di questi corsi d'acqua con matrice sabbiosa. Cappello di piccole dimensioni 1,5-2 cm di diametro color bruno chiaro, più chiaro perifericamente e striato per trasparenza fino a un terzo del raggio, presenza di un piccolo umbone, lamelle da ocra chiaro nei giovani esemplari fino brunatre in maturazione, da adnate ad appena decorrenti sul gambo, quest'ultimo si presenta cilindrico, appena bulbosetto alla base, un poco più scuro del cappello e percorso da fibrille longitudinali più chiare, più evidenti verso l'apice, consistente, non fragile, lungo almeno il doppio del cappello, sapore un poco sgradevole, odore leggero, rafanoide-terroso, difficile da identificare per la crescita su lettiera umida. Regione Lombardia, pianura Bergamasca, loc. Spirano; Novembre 2011; Foto, commento e microscopia di Massimo Biraghi. Spore (10,45)10,74-11,85-12,60(12,62) × (4,82)5,02-5,2-5,78(5,99) µm; Q = (1,99)1,99 - 1,99 - 2,351(2,47); da lungamente agmidaliformi a fusiformi con restrigimento basale, presenza di apicolo, marcatamente verrucose, appena destrinoidi. Osservazione in Acqua. Osservazione in Rosso Congo. Osservazione in Melzer. Basidi lunghi 30-40 µm, banali, tetrasporici, ma anche bisporici, con GAF. Cheilocistidi abbondanti lunghi fno a 60 µm, con base ventricosa e con collo stretto e allungato, non capitulato. Osservazione 400×. Osservazione 1000×. Caulocistidi presenti verso l'apice del gambo, simili ai cheilocistidi. Osservazione 400×. Osservazione 1000×. GAF presenti in tutte le strutture osservate, cutis composta da ife cilindriche a ovoidi, alcune con pigmento incrostante. -
Naucoria salicis P.D. Orton 1960
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Naucoria salicis P.D. Orton 1960 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Sinonimi Naucoria macrospora J.E. Lange 1940 Alnicola macrospora J.E. Lange ex J. Favre 1948 Alnicola langei Kühner 1957 Naucoria langei Kühner 1957 Alnicola salicis (P.D. Orton) Bon 1979 Foto e Descrizioni Ritrovamento in ambiente molto umido in prossimità di Carpino e nelle vicinanze di Castagno, colline bergamasche 400 m s.l.m. L'odore rafanoide con timbri di cacao e le lamelle crema, ocra chiaro con riflessi rosa, potrebbero far pensare a un Hebeloma sp. Analizzando i caratteri morfocromatici, in particolare le striature al margine del cappello, l'aspetto in generale esile a confronto di Hebeloma sp., le colorazioni delle lamelle da giovane con riflessi rosati, il gambo sericeo e lucente, percorso logitudinalmente da striature, nonchè le colorazioni pileiche "monotone" che evocavano Entoloma sp., indirizzano correttamente verso il Genere Naucoria. Le specie del Genere Naucoria (= Alnicola) crescono abitualmente sotto Alnus spp., ma in alcuni casi, come questo, il ritrovamento è stato effettuato sotto Carpino e Castagno, con Quercia poco distante. L'analisi microscopica conferma questa ipotesi. La Sez. Salicicolae Kühner è caratterizzata da assenza di GAF nei tessuti osservati e forma particolare dei cheilocistidi. In questa sezione si distingue una sola specie con misure delle spore che concordano con i rilevamenti microscopici osservati. Cheilocistidi 35-45 × 5-8 µm, cilidrici, leggermenti clavati, subcapitulati e con apice dilatato fino a 12 µm. Spore (14)15-20(22) × 8-9 (10) µm. Basidi bisporici 25 × 8 µm. Regione Lombardia; Maggio 2010; Foto, descrizione e microscopia di Massimo Biraghi. Spore (14)15-20(22) × 8-9(10) µm, da largamente amigdaliformi a lungamente ellissoidali o subfusiformi, grossolanamente verrucose. Basidi banali, bisporici 25 × 8 µm. Cheilocistidi numerosi 35-45 × 5-8 µm, da subcilindrici a leggermente clavati, subcapitulati, con apice dilatato e largo fino a 12 µm. Pileipellis. -
Naucoria escharioides (Fr. : Fr.) P. Kumm. 1871
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Naucoria escharioides (Fr. : Fr.) P. Kumm. 1871 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Strophariaceae Foto e descrizione Cappello 1-3 cm, ricoperto da resti di velo biancastri che lasciano intravedere un colore ocra chiaro sottostante, più scuro al centro, fino a brunastro ed in prossimità di un evidente umbone, lamelle di un bel ocra-giallastro carico, gambo cilindrico, subconcolore, più scuro verso la base, imbrunente alla manipolazione. Carne esigua, fragile. odore sub nullo o leggermente rafanoide, sapore non amaro o non rilevante. Microscopia Basidi tetrasporici. Cheilocistidi urticoidi-subfusiformi. Spore amigdaliformi, rugose, con verruche. 60 misurazioni N° Long. Larg. Q Moy 9,30 5,07 1,84 Min 8,26 4,50 1,67 Max 10,35 5,63 2,18 Media 9,23 5,10 1,82 Long.: N = 32; dMd ; (9,23)9,489-9,92-11,527(12,13) Larg.: N = 32; dMd ; (4,84)4,939-5,365-5,987(6,47) Q: N = 32; dMd ; (0,30)1,76-1,80714285714286-2,057(2,07) N° Long. Larg. Q Moy 10,28 5,48 1,83 Min 9,23 4,84 Max 12,13 6,47 2,07 Media 10,19 5,47 1,86 Lxl: N = 32; dMd; (9,23)9,49-9,92-11,53(12,13) × (4,84)4,94-5,365-5,99(6,47); Qm = 1,83. Regione Lombardia; Novembre 2013; Foto, descrizione e microscopia di Massimo Biraghi. Microscopia Basidi tetrasporici. Cheilocistidi urticoidi-subfusiformi. Spore amigdaliformi, rugose, con verruche osservazione in rosso congo 100×. Spore acqua 100×. -
Myxomphalia maura (Fr. : Fr.) Hora 1960
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Myxomphalia maura (Fr. : Fr.) Hora; Regione Campania; Ottobre 2008; Foto di Felice Di Palma. Microscopia Amiloidia delle spore. Spore e relativa misura. Cheilocistidi. Cheilocistidi e basidi. -
Myxomphalia maura (Fr. : Fr.) Hora 1960
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Myxomphalia maura (Fr. : Fr.) Hora; Regione Lazio; Febbraio 2008; Foto di Mauro Cittadini. -
Myxomphalia maura (Fr. : Fr.) Hora 1960
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Myxomphalia maura (Fr. : Fr.) Hora; Regione Sardegna; Novembre 2007; Foto di Pietro Curti.
