
Archivio Micologico
-
Numero contenuti
15665 -
Iscritto
-
Ultima visita
Tipo di contenuto
Profili
Forum
Orchidee
Diventa Socio
Calendario
Tutti i contenuti di Archivio Micologico
-

Morchella esculenta (L. : Fr.) Pers. 1794
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Morchella esculenta (L. : Fr.) Pers.; Regione Umbria; Aprile 2012; Foto di Mario Iannotti.
-
Agaricus braendlei L.A. Parra & M.M. Gómez 2018
Archivio Micologico ha postato un topic nell'area Funghi Commestibili
Agaricus braendlei L.A. Parra & M.M. Gómez 2018 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Agaricaceae Genere Agaricus Sottogenere Agaricus Sezione Agaricus Sinonimi Agaricus argenteus Braendle ex Peck 1899. Etimologia L'epiteto Agaricus deriva dal greco αγαρικόν = fungo degli Agari; perché secondo Dioscoride proveniva dall'Ucraina, che a quel tempo veniva chiamata "Terra di Agaria". L'epiteto braendlei è dedicato a F.J. Braendle, che Peck ha indicato come primo scopritore della specie. Cappello 3-7 cm, emisferico, poi da convesso ad appianato; margine intero oppure, a volte, appendicolato per i residui del velo parziale. Superficie asciutta, secca. Colore da biancastro a grigio argenteo. KOH sul cappello negativo. Lamelle Fitte, libere al gambo, rosa pallido che scuriscono con l’età, fino a diventare bruno-nerastre. Filo lamellare concolore. Gambo 3-6 cm e spesso 1-1,5 cm, cilindrico assottigliato verso la base appuntita. Biancastro, imbrunente verso la base. Anello Il velo sottile, fragile, può lasciare traccia nei giovani esemplari sul margine del cappello e forma un anello bianco, molto evanescente, spesso ridotto a semplici resti sulla parte alta del gambo. Micelio basale di colere bianco. Carne Soda, bianca. Odore e sapore non significativi. Habitat In primavera e autunno, tra l’erba delle radure di terreni compatti e incolti, in parchi, sempre fuori dai boschi. Microscopia Spore (6,5)6,7-8,0(8,45) × (4,6)4,8-5,5(5,6) µm; Q = (1,2)1,3-1,5(1,6); N = 30; ellittiche, lisce, bruno nerastre in massa. Basidi tetrasporici. Cheilocistidi assenti. Commestibilità e Tossicità Commestibile. La specie appartiene agli Agaricus senza odori fenolici e ingiallimenti alla corrosione, né rizoidi alla base del gambo. Non sono conosciuti micetismi attribuibili a questa specie. Specie simili Agaricus campestris è la specie più simile, al punto che Kerrigan, uno dei maggiori studiosi del genere Agaricus, ritiene che molte raccolte Nord Americane etichettate come Agaricus campestris debbano invece essere ascritte ad Agaricus braendlei, il suo sosia Nord Americano Orientale. In effetti le due specie si differenziano semplicemente per la dimensione delle spore, maggiori o minori di 8 µm di lunghezza, e per la diversa filogenesi molecolare. Altri elementi distintivi rispetto alla specie qui citata sono: il contesto al taglio, bianco immutabile, mentre in Agaricus campestris la carne diventa leggermente rosata nel cappello e nella parte superiore del gambo; una fascia imbrunente verso la base del gambo che in Agaricus campestris manca; odore insignificante contro quello fungino di Agaricus campestris. Note nomenclaturali Agaricus argenteus Braendle ex Peck 1899, nome con il quale fino al 2018 è stata chiamata la specie in esame, è considerato un nome illegittimo, in quanto sinonimo posteriore di Agaricus argenteus Whit. 1792. William Whitering, un medico e botanico Britannico vissuto nel 1700, scopritore, tra le altre, di talune applicazioni mediche della Digitalis, scrisse tra il 1741 ed il 1799 un'opera in tre volumi: "A Botanical Arrangement of British Plants"; nel terzo volume vengono trattati anche i Funghi, ed in particolare a pagina 354 viene descritto Agaricus argenteus come un fungo con: "Cappello marrone chiaro acquoso, con il centro più scuro, con cuticola farinosa, striata al margine da giovane, rugosa ed intrecciata da vecchio; lamelle bianco-brunastro acquoso, con il filo argenteo brillante (da cui il nome), che cresce a grappoli sotto Olmo". Difficile dire a quale fungo si volesse riferire dato che, tra l'altro, mancano disegni e rappresentazioni delle specie descritte. Bibliografia LIU, A., DAI, R., ZHANG, M., CAO, B., XI, Y., WEI, S. & ZHAO, R., 2020. Species of Agaricus section Agaricus from China. Phytotaxa 452 (1): 001–018. PARRA, L.A., CAPPELLI, A., KERRIGAN, R.W., BIZIO, E., 2018. Agaricus porphyrocephalus subsp. alpinus a new subspecies collected in the Italian Alps. Micologia e Vegetazione Mediterranea, 33(2): 67-68. WHITERING, W., (1741-1799). A Botanical Arrangement of British Plant. Vol. 3: 354. Scheda di proprietà AMINT realizzata da Felice Di Palma - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Sardegna; Novembre 2007; Foto di Felice Di Palma. Gli esemplari rappresentati in foto, inizialmente determinati come Agaricus campestris, sono stati oggetto di analisi molecolare. L'operazione si è rivelata non agevole, probabilmente per il cattivo stato di conservazione degli esemplari raccolti, con ben due tentativi di estrazione andati a vuoto. Quando siamo finalmente riusciti ad estrarre una sequenza, grazie ai preziosi consigli dell'amico Valerio Mazzella, Ricercatore presso il Dipartimento di Ecologia Marina Integrata della Stazione Zoologica di Napoli, questa si presentava comunque molto rovinata e non in grado di fornire soluzioni risolutive. Solo l'intervento paziente di Luis Alberto Parra, il massimo esperto Europeo del genere Agaricus che grazie alla sua notevole esperienza è riuscito a "pulire" adeguatamente la sequenza, ha permesso l'ascrizione alla specie qui descritta del nostro ritrovamento. Spore (6,5)6,7-8,0(8,45) × (4,6)4,8 - 5,5(5,6) µm; Q = (1,2)1,3-1,5(1,6); N = 30. Osservazione in acqua, a 1000×. Filo lamellare fertile, basidi e spore. Osservazione in Rosso Congo, a 1000×. -
Coprinopsis marcescibilis (Britzelm.) Örstadius & E. Larss. 2008
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Coprinopsis marcescibilis (Britzelm.) Örstadius & E. Larss. 2008 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Psathyrellaceae Sinonimi Psathyrella marcescibilis (Britzelm.) Singer 1951 Psathyrella fragilissima (Kauffman) A.H. Sm. 1972 Psathyrella lactea (J.E. Lange) J.E. Lange 1940 Hypholoma marcescibile (Britzelm.) Sacc. 1895 Etimologia L'epiteto Coprinopsis deriva dal genere “Coprinus” con il suffisso - ópsis dal greco "ὄψις" [ópsis] = dall'aspetto di, per l'aspetto simile a Coprinus. L'epiteto marcescibilis deriva dal latino "marcésco" iniziare a marcire, marcescibile, che marcisce facilmente. Cappello Pileo convesso di color nocciola rossiccio, igrofano, pertanto il carpoforo assume tonalità diverse in base all’umidità, da nocciola brunastro a beige chiaro quando si asciuga, inizialmente umbonato, con depressione piatta e discolore in corrispondenza del disco in soggetti maturi, margine striato, eccedente, con evidenti fiocchi di velo dentati e triangolari. Lamelle Imenoforo a lamelle regolari, leggermente panciute, alternate a lamellule di ogni dimensione, filo lamellare discolore biancastro finemente fimbriato. Anello Filamentoso, fugace, ben evidente nei soggetti giovani a proteggere l'imenoforo. Gambo Stipite 10-12 × 0,5-0,7 cm, allungato, cilindrico o sinuoso, striato all'inserzione con l'imenoforo, cavo all'interno, ricco di fiocchi farinosi su tutta la lunghezza. Carne Poco consistente, priva di odore e sapore particolare. Habitat Fungo saprotrofo, vegeta su detriti organici e fogliame sia di aghifoglia che latifoglia. Microscopia Cheilocistidi utriformi, con peduncolo basale. Pleurocistidi non osservati. Trama lamellare costituita da cellule globose. Giunti a fibbia presenti in tutti i tessuti. Pileipellis mista, costituita da un sottile strato esterno di ife parallele, strette e allungate e da uno strato inferiore formato da cellule globose e ellissoidali, con setti ristretti. Basidi clavati, tetrasporici. Spore (9,8) 10,5-11,6 (12,1) × (5,8) 6,2-7,2 (7,8) µm; Q = (1,5) 1,52-1,8 (1,9); N = 50; Media = 11,1 × 6,7 µm; Qm = 1,7; amigdaliformi in vista laterale, ovoidali in vista frontale, appiattite in corrispondenza del poro germinativo, lisce, di colore bruno scuro, quasi nero, con apicolo evidente. Stipitipellis costituita da cellule allungate, cilindriche. Caulocistidi di forma e dimensione variabili, cilindracei, sinuosi, subcapitulati, contenenti pigmento ocraceo, in parte disciolto, in parte contenuto in guttule. Commestibilità e tossicità Non commestibile. Specie simili Coprinopsis marcescibilis morfologicamente e per il portamento, potrebbe essere confusa con altre specie del genere Coprinopsis e genere Psathyrella. Solo un attento esame dei caratteri microscopici, contribuisce ad una corretta determinazione. Coprinopsis pseudomarcescibilis Heykoop, G. Moreno & P. Alvarado è una specie sosia, difficile da separare morfologicamente, si differenzia per la taglia più piccola e microscopicamente per le spore che sono più lunghe, con valori di 13,3-14,5 μm. Coprinopsis musae Örstadius & E. Larss carpofori di piccola taglia, con spore piccole, molto pallide, valori di 9-10,5 × 5,8-6,2 μm. Psathyrella candolleana (Fr.: Fr.) Maire si differenzia morfologicamente per le colorazioni più tenui, per la taglia dei carpofori meno slanciata, microscopicamente per le spore piccole valori di 6-8 × 4-5 µm e per la pileipellis formata da un'unico strato di cellule subglobose. Bibliografia HEYKOOP, M., MORENO, G. & ALVARADO, P., 2017. Coprinopsis pseudomarcescibilis Heykoop, G. Moreno & P. Alvarado, sp. nov. Persoonia 38, Fungal planet description sheet 594: 320-321. ÖRSTADIUS, L., RYBERG, M. & LARSOON, E., 2015. Molecular phylogenetics and taxonomy in Psathyrellaceae (Agaricales) with focus on psathyrelloid species: introduction of three new genera and 18 new species. Mycol Progress 14, 25. Doi https://doi.org/10.1007/s11557-015-1047-x. Doi AA.VV., 2012. Funga Nordica. Agaricoid, boletoid and cyphelloid genera. Ed. Nordsvamp. WÄCHTER, D. & MELZER, A., 2020. Proposal for a subdivision of the family Psathyrellaceae based on a taxon-rich phylogenetic analysis with iterative multigene guide tree. Mycological Progress 19: 1151-1265. DOI https://doi.org/10.1007/s11557-020-01606-3. Scheda di proprietà AMINT realizzata da Giovanni Galeotti - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Umbria; Dicembre 2021; Foto e microscopia di Giovanni Galeotti. (exsiccatum GG211229-01) Residui velari sul margine, anello dissociato e filamentoso nel soggetto giovane. Stipite striato all'inserzione con l'imenoforo, filo lamellare discolore. I soggetti osservati erano disseminati su una zona piuttosto vasta, con crescita isolata su detriti organici di aghifoglia mista a latifoglia. Zona discale depressa e discolore, in corrispondenza dell'inserzione al pileo dello stipite. Trama lamellare costituita da cellule globose, evidenti cheilocistidi sulla destra della foto. Osservazione in Rosso Congo, a 40×. Cheilocistidi utriformi. Osservazione in Rosso Congo, a 400×. Caulocistiti con GAF. Osservazione in Rosso Congo, a 400×. Caulocistidi e GAF, con pigmento ocraceo, contenuto in guttule, e in parte disciolto. Osservazione in Rosso Congo, a 1000×. Velo parziale prelevato sul margine del pileo. Osservazione in Rosso Congo, a 100×. Ife del velo. Osservazione in Rosso Congo, a 400×. Sezione della cuticola. Osservazione in Rosso Congo, a 100×. Cuticola mista, costituita, da un sottile strato ife parallele, strette e allungate, e da uno strato inferiore, formata da cellule globose e ellissoidali, con setti ristretti. Osservazione in Rosso Congo, a 400×. Basidi clavati, tetrasporici. Osservazione in Rosso Congo, a 400×. Spore (9,8) 10,5-11,6 (12,1) × (5,8) 6,2-7,2 (7,8) µm; Q = (1,5) 1,52-1,8 (1,9); N = 50; Media = 11,1 × 6,7 µm; Qm = 1,7. Spore bruno nerastre, amigdaliformi in vista laterale, ovoidali in vista frontale. Osservazione in Rosso Congo a 1000×.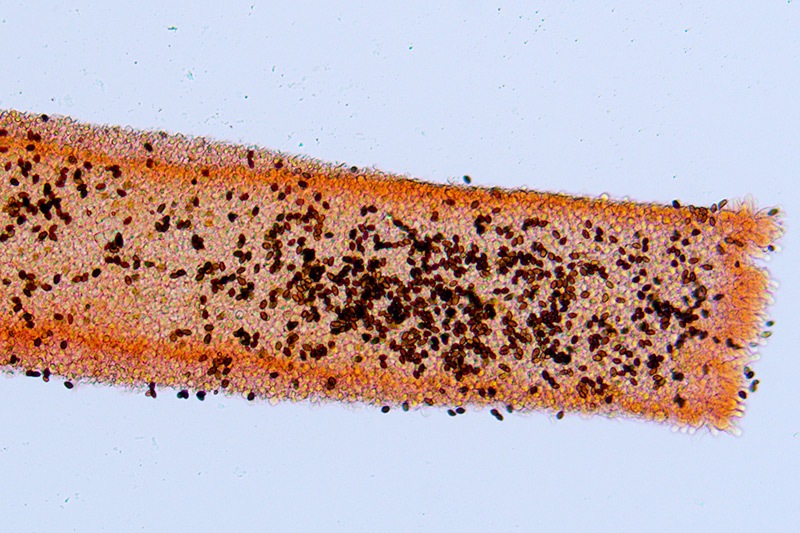
-
Aleuria luteonitens (Berk. & Broome) Gillet 1886
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Aleuria luteonitens (Berk. & Broome) Gillet 1886 Tassonomia Divisione Ascomycota Classe Pezizomycetes Ordine Pezizales Famiglia Pyronemataceae Sinonimi Peziza luteonitens Berk. & Broome 1851 Otidea luteonitens (Berk. & Broome) Massee 1895 Etimologia L'epiteto Aleuria deriva dal greco ἄλευρον [áleuron] = farina, per l'apotecio che esternamente è forforaceo. L'epiteto Luteonitens deriva dal latino lúteus = giallo, e nitens = splendente, lucente, di un giallo brillante. Ascoma Apotecio sessile, 1-1,5 cm di diametro, con il margine finemente crenulato, di colore giallo vivo, a forma di coppa discoide, a volte disteso, l’imenio è liscio e concolore. La superficie esterna, sempre su tonalità giallastre presenta forfora e pruinosità. Fungo gregario e cespito, fruttifica in gruppetti, con soggetti che si presentano appressati tra loro. Carne Carne chiara, giallognola e acquosa, non geme latice alla frattura, priva di odore. Habitat Nel bosco in ambiente umido, su terreno sciolto o argilloso, con presenza di detriti organici e muschio. Microscopia Spore (10,1) 10,4-11,8 (12,7) × (5,6) 5,9-6,7 (7,1); Q = (1,5) 1,6-1,9 (2,0); N = 31; Media = 11,0 × 6,3; Qm = 1,8; ellissoidali ornate da sottili verruche, che formano uno pseudoreticolo incompleto, escluse le asperità, che misurano µm 1-1,5. Aschi (147,9) 156,0-193,4 (197,8) × (7,5) 8,5-9,9 (10,2) µm; Q = (17,1) 17,3-20,4 (20,5); N = 11; Media = 170,6 × 9,0 µm; Qm = 18,8; uniseriati ottasporici, opercolati, pleurorinchi, non amiloidi. Parafisi cilindriche con lunghezza maggiore degli aschi, con apice dilatato e ricurvo, contengo un pigmento ocraceo disperso. Excipulum ectale formato da cellule globose. Peli sulla superficie esterna formati da corte cellule cilindriche, arrotondate, con strozzature in corrispondenza dei setti, ultima cellula con estremità arrotondata. Excipulum medullare a textura intricata, formata de cellule distese orizzontalmente, catenulate. Commestibilità e tossicità Non commestibile. Specie simili Aleuria cestrica (Ellis & Everh.) Seaver è certamente la specie sosia, simile morfologicamente, gialla ma con sfumature aranciate, si presenta anche su terreno nudo e microscopicamene, si differenzia per le spore più piccole 8-10 x 5-6 µm. Aleuria bicucullata Boud. ha spore generalmente più larghe 10-12 x 6-7,5 µm. provviste di nervature, che formano un reticolo elicoidale in rilievo, con creste alte fino a 4-4,5 µm. Aleuria aurantia (Pers. : Fr.) Fuckel differisce per dimensioni più grandi e per tonalità aranciate dell'apotecio e per le spore decisamente più grandi 18-20,5 x 11-12 µm. Osservazioni Auleria luteonitens si riconosce, tra le specie appartenenti a questo genere, per i colori giallo brillanti dell'apotecio, per le piccole dimensioni. Microscopicamente per le dimensioni delle spore, che presentano verruche sottili, interconnesse a formare delle creste che convergono a costituire uno pseudoreticolo incompleto. Bibliografia BILLEKENS, P., 1994. Aleuria bicucullata versus Aleuria luteonitens. Karstenia. MEDARDI, G., 2006. Atlante fotografico degli Ascomiceti d'Italia. Ed. AMB. VAN VOOREN, N., 2014. Cahiers de la FMBDS 4: 4-5. Scheda di proprietà AMINT realizzata da Giovanni Galeotti - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Umbria; Ottobre 2021, Foto e microscopia di Giovanni Galeotti. (Exsiccatum GG211008-01) Soggetti rinvenuti su muschio, in un bosco planiziale altitudine 400 metri s.l.m. tagliato di recente, con prevalenza di Quercus cerris e Populus alba, fruttifica formando dei circoli. Margine dell'apotecio più chiaro e finemente crenulato. Soggetti con colorazione giallo-aranciato. Sezione dell'apotecio. Osservazione in Rosso Congo, a 40×. Excipulum ectale. Osservazione in Rosso Congo a 100×. Aschi non amiloidi. Ossarvazione in Melzer a 400×. Excipulum medullare. Osservazione in Rosso Congo a 400×. Peli sulla superficie esterna formati da corte cellule cilindriche, arrotondate, con strozzature in corrispondenza dei setti, ultima cellula con estremità arrotondata. Osservazione in Rosso Congo a 400×. Aschi e parafisi. Osservazione in Rosso Congo, a 400×. Parafisi settate, spesso ricurve e dilatate all'apice. Osservazione in Rosso Congo, a 400×. Aschi e parafisi. Osservazione in Rosso Congo, a 1000×. Spore. (10,1) 10,4-11,8 (12,7) × (5,6) 5,9-6,7 (7,1); Q = (1,5) 1,6-1,9 (2,0); N = 31; Media = 11,0 × 6,3 ; Qm = 1,8 Osservazione in acqua a 1000×. -
Warcupia cupulata M. Carbone, Galeotti, Lezzi, Athanasiadis & P. Alvarado 2021 Tassonomia Divisione Ascomycota Classe Pezizomycetes Ordine Pezizales Famiglia Otideaceae Etimologia L’epiteto di genere Warcupia è dedicato al Dr. Jack H. Warcup. L’epiteto cupulata deriva dal Latino cupulatus = cupolato, per la forma a coppa dell'apotecio. Ascoma L’apotecio si presenta con un diametro di 5-30 mm, alto 20-40 mm, è prevalentemente intero, anche se alcuni campioni presentano un'apertura laterale (come avviene tipicamente nel genere Otidea). La superficie esterna è pubescente, pruinosa e furfuracea, di color nocciola chiaro con riflessi aranciati da giovane ed è più scura a completa maturità. Il margine è finemente crenulato negli esemplari giovani, poi denticolato e decorato da verruche color ruggine a maturità. L’imenio si presenta liscio, subconcolore alla superficie esterna, con riflessi grigiastri in gioventù, grinzoso. Il fungo si presenta gregario o fascicolato. Stipite Lo stipite è ben distinto, sviluppato e consistente, concolore con l’apotecio, pieno alla sezione, rivestito da una pruina biancastra, dilatato all’inserzione della base dell’apotecio. Il micelio basale è biancastro. Carne La carne si presenta di consistenza ceracea, non presenta odore rilevante, né sapore particolare. Habitat Al momento risultano soltanto quattro località di rinvenimento di Warcupia cupulata. Tutte le raccolte segnalate hanno come carattere comune la presenza di Cupressus spp. I ritrovamenti umbri del 2019, 2020 e 2021, compreso il ritrovamento dell’olotipo, provengono dalla provincia di Perugia, da un parco situato a 400 m s.l.m. I campioni sono stati trovati su substrato ricco di humus e detriti organici. Nella zona dove sono avvenuti questi ritrovamenti, sono presenti molte specie arboree, di cui alcune alloctone e di particolare pregio. A breve distanza tra di loro sono presenti: Cupressus lusitanica, C. macrocarpa, Quercus ilex, Picea orientalis, P. abies, Abies cephalonica, A. alba, A. pinsapo, Taxus baccata e Calocedrus decurrens. Una seconda località di ritrovamento si trova in Grecia, dove gli esemplari sono stati rinvenuti in un bosco di conifere con Pinus brutia e Cupressus sempervirens f. horizontalis. Un terzo ritrovamento è stato segnalato in Italia, nella regione Toscana, sotto Cupressus macrocarpa; e un quarto in Spagna, sotto Eucalyptus sp. e Cupressus sp. Microscopia Ascospore (8,8-)9,5-11(-11,8) × (4,9-)5,6-6,7(-7,3) μm; Q = (1,4-)1,5- 1,9(-2,0); media = 10,3 × 6,1 μm; Qm = 1,7 (n = 161), lisce, ialine, da ellissoidi a leggermente ovoidali, biguttulate, con parete fino a 0,4 μm di spessore. Parafisi cilindriche, settate, ramificate alla base, leggermente più lunghe degli aschi, con apici dilatati e piegati a uncino, larghe fino a 2,5 μm. Aschi (113,6-)129,5-152,4 (-227,4) × (6,7-)7,8-9,3(-9,5) μm, Q = (12,7-)14,8-19,5(-26,3), media = 144,6 × 8,4 μm, Qm = 17,3 (n = 30); cilindrici, non amiloidi, opercolati, pleurorinchi, contengono 8 spore uniseriate. Subimenio si presenta come una sottile zona più scura, composta da cellule piuttosto fitte. Excipulum medullare composto da cellule disposte con textura intricata, formato da ife (6,8-)8,1-13(-17,5) μm di larghezza, media = 10,4 μm (n = 30), ialine, a parete sottile. Excipulum ectale si presenta con textura globulosa angolare nella parte interna, con cellule (8,7-)9,6-13,7(-16,2) μm diam., media = 10,4 μm (n = 30), moderatamente spesse, da ialine a giallastre. La superficie esterna dell'ascoma è coperta da verruche formate da cellule che presentano una textura da globulosa a globuloso-angolare. Micelio basale formato da ife larghe 8,5 μm, parete fino a 1 μm di spessore, ialine, settate, a parete liscia. Specie simili Al genere Warcupia fino a poco tempo fa apparteneva una sola specie cleistotecioide: Warcupia terrestre, vedi Paden et al. (1971). La specie Warcupia cupulata presenta invece apoteci ben formati, di discrete dimensioni e con spore più piccole. Alcuni esami molecolari basati su marcatori ITS, LSU, tef1 e rpb2 hanno dimostrato come le due specie, nonostante un aspetto macroscpico profondamente differente, appartengano allo stesso clade, come spiegato in dettaglio in Carbone et al. (2021). Warcupia cupulata, sia per microscopia che per forma degli apoteci può ricordare: le specie a coppa intera del genere Otidea, da cui si differenzia per avere ascospore più piccole; le specie del genere Arpinia da cui si differenzia per le estremità delle parafisi ricurve; le specie del genere Tarzetta da cui si differenzia per gli aschi pleurorinchi. Commestibilità e tossicità Attualmente non è conosciuta la commestibilità di questa specie. Nota Per informazioni più dettagliate fare riferimento all'articolo in cui la specie è stata pubblicata: CARBONE M., GALEOTTI, G., LEZZI, T., ATHANASIADIS, A. & ALVARADO, P., 2021. Warcupia cupulata a new cup-shaped species in the cleistothecioid genus Warcupia (Otideaceae, Pezizales). Ascomycete.org, 13 (4): 145-156. Bibliografia CARBONE M., GALEOTTI, G., LEZZI, T., ATHANASIADIS, A. & ALVARADO, P., 2021. Warcupia cupulata a new cup-shaped species in the cleistothecioid genus Warcupia (Otideaceae, Pezizales). Ascomycete.org, 13 (4): 145-156. PADEN, J.W. & CAMERON, J.V., 1971. Morphology of Warcupia terrestris, a new ascomycete genus and species from soil. Canadian Journal of Botany 50 (5): 999-1001. Scheda di proprietà AMINT realizzata da Giovanni Galeotti e Tomaso lezzi - Approvata e Revisionata dal CLR Micologico di AMINT. Si ringrazia Matteo Carbone, autore principale dell'articolo Carbone et al. (2021), per la revisione del testo. Regione Umbria; Dicembre 2019, Dicembre 2020, Gennaio 2021; Foto di Giovanni Galeotti e Tomaso Lezzi. [Exsiccata TUR-A 209463, TUR-A 209464 (Paratypi); TUR-A 209465 (Holotypus)] Giovani soggetti con superficie esterna pruinosa, e tonalità nocciola aranciate. Sezione dello stipite. Si evidenziano le tonalità brunastre degli apoteci, con sfumature bruno violacee grinzose dell'imenoforo, in soggetti molto maturi Esemplari con il margine dell'apotecio fortemente denticolato, in soggetti adulti in piena fase di sviluppo. Sezione dell'apotecio. Osservazione in Rosso Congo, a 40×. Sezione dell'apotecio. Osservazione in Rosso Congo, a 100×. Aschi e parafisi. Osservazione in Rosso Congo, a 400×. Aschi e parafisi. Osservazione in Rosso Congo, a 400×. Parafisi settate, ricurve all'apice. Osservazione in Rosso Congo, a 400×. Aschi ottasporici, uniseriati, pleurorinchi. Osservazione in Rosso Congo, a 400×. Aschi osservati in acqua, a 400×. Aschi e parafisi. Osservazione in Melzer a 400x. Spore. Osservazione in Rosso Congo, a 1000×. Spore. Osservazione Acqua, a 1000×. Spore. Osservazione Blu cotone, a 1000×. Excipulum ectale formato da cellule globose, globose angolari. Osservazione in Rosso Congo, a 400×. Excipulum ectale. Osservazione in Rosso Congo, a 400×. Excipulum medullare a texura intricata. Osservazione in Rosso Congo, a 400×.
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Luglio 2015; Foto di Francesco Tozzi. Esemplari con queste colorazioni venivano qualche tempo fa ascritti a Boletus luridiformis var. discolor (Quél.) Krieglst.
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Liguria, Sassello, raduno AMINT; Ottobre 2012; Foto di Tomaso Lezzi. Boletaceae a pori rossi, come si vede bene dalla foto macro il rosso si trova solo sul bordo dei pori, non sui tubuli, che sono gialli. Netto viraggio al blu scuro al tocco.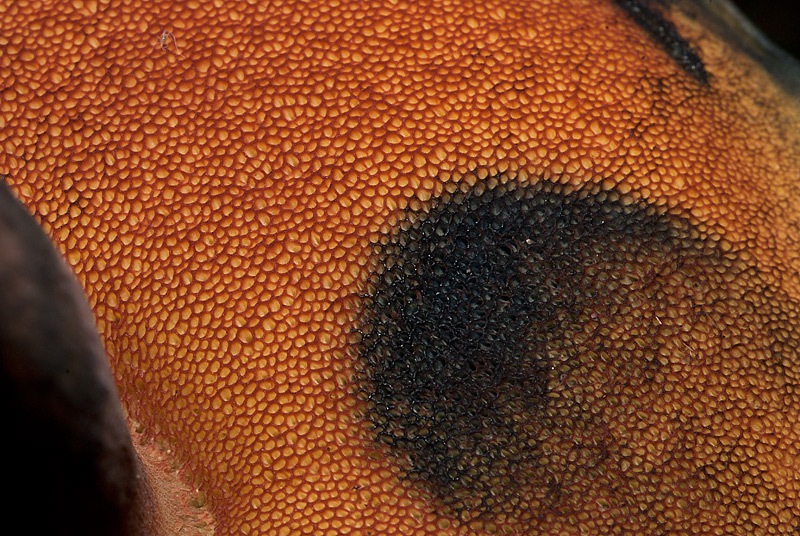
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Lazio; Luglio 2011; Foto di Tomaso Lezzi. Boletaceae a pori rossi, cappello marrone vellutato, gambo senza reticolo coperto da punteggiatura rossa, forte viraggio al taglio e alla contusione; qui in una versione solo apparentemente lignicola, perché cresceva incastrato in un tronco.

-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Lazio; Luglio 2015; Foto di Francesco Tozzi.


-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Umbria; Luglio 2010; Foto di Luigi Minciarelli. Appennino umbro-marchigiano, versante umbro, al bordo di faggeta. Sezione. Foto di Tomaso Lezzi. La sezione con il viraggio al blu. Un particolare dei pori rossi, che diventano blu al tocco. Un particolare della puntinatura rossa sul gambo.



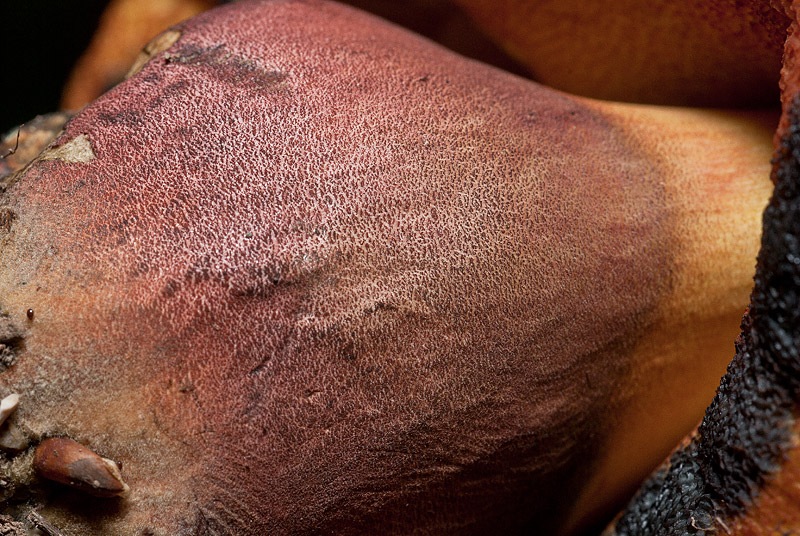
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Calabria; Luglio 2009; Foto di Pietro Curti.
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Piemonte, Valsesia, Scopa (VC); Settembre 2011; Foto di Alessandro Remorini. Ritrovamento effettuato a 650 m s.l.m.

-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Lombardia, colline bergamasche, località Palazzago; Ottobre 2007; Foto di Massimo Mantovani.
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Calabria; Settembre 2005; Foto di Antonio Lupo.
-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini; Regione Liguria; Ottobre 2009; Foto di Alessandro Remorini.

-
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014
Archivio Micologico ha postato un topic nell'area Funghi Commestibili
Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini 2014 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Boletales Famiglia Boletaceae Nome italiano Boleto dal piede rosso. Sinonimi Boletus erythropus Pers. : Fr. 1796 Boletus luridiformis Rostk. 1844 Neoboletus erythropus (Pers. : Fr.) C. Hahn 2015 Sutorius luridiformis (Rostk.) G. Wu & Zhu L. Yang 2016 Etimologia Dal latino luridiformis = che ricorda Boletus luridus, dal latino fòrma = forma, foggia e dal termine micologico luridus. Cappello 10-20(25) cm, emisferico poi convesso ed infine appianato, tomentoso, con cuticola finemente vellutata, asciutta, di colore bruno scuro, talvolta bruno-rossiccio, con sfumature più chiare, più facilmente verso il margine; orlo regolare e leggermente eccedente. Tuboli e pori Tuboli lunghi, arrotondati al gambo, quasi liberi, gialli poi olivastri, al taglio rapidamente viranti al blu; pori piccoli, rotondi, giallo-arancio nei soggetti molto giovani, poi da rosso-arancio a rosso sangue, generalmente decolorati al margine, imbluenti alla pressione. Gambo 7-15 × 3-7 cm, robusto, da obeso a cilindraceo-ventricoso, dilatato verso la base generalmente arrotondata, ricoperto da una fine punteggiatura rosso sangue su fondo giallastro, giallo-arancio, rosso più o meno scuro alla base e imbluente alla manipolazione. Carne Soda e compatta nel giovane, tardivamente molliccia nel cappello, dura nel gambo, generalmente giallastra-giallo cromo, talvolta rossastra alla base, virante al blu in maniera intensa al taglio; odore fruttato, sapore dolce. Habitat Relativamente diffuso dall’estate all’autunno, a quote che variano tra 200-1000 m s.l.m., sempre sotto latifoglia in ambiente mesofilo, almeno secondo gli ultimi studi molecolari. I ritrovamenti di “Boletus erythropus” sotto latifoglie (Fagus sylvatica, Castanea sativa, Quercus sspp.), secondo alcuni autori sarebbero da attribuire a Neoboletus xanthopus (Klofac & A. Urb.) Klofac & A. Urb. = Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini. Ultimamente Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizzini viene anche proposto da taluni studiosi con il current name di Sutorius luridiformis (Rostk.) G. Wu & Zhu L. Yang 2016. Microscopia Spore bruno-olivastre in massa, ellissoidali, lisce, 12-16 × 5-6 µm. Commestibilità o Tossicità Commestibile dopo adeguata cottura che consente di eliminare alcune tossine termolabili. Osservazioni Per i cromatismi scuri del cappello e la cuticola vellutata viene spesso confuso al primo frettoloso sguardo con i Boletus della sez. edules. Specie simili Neoboletus praestigiator (R. Schulz) Svetash., Gelardi, Simonini & Vizzini è a tutti gli effetti un suo sosia, e si distingue, oltre che attraverso gli esami molecolari, per la crescita sotto aghifoglie e latifoglie nella fascia subalpina. Viene spesso confuso soprattutto con Suillelus luridus (Schaeff. : Fr.) Murrill = Boletus luridus Schaeff. : Fr. il quale si distingue per la cuticola vellutata e non di aspetto “lurido”, il gambo punteggiato e non con un reticolo marcato e per il sub imenio giallastro e mai arancione-rossastro. Neoboletus junquilleus (Quél.) Gelardi, Simonini & Vizzini = Boletus erythropus var. junquilleus (Quél.) Bon, è praticamente un suo sosia con cappello, imenio (tubuli e pori) e gambo giallastri, mai con tonalità rosse. Suillellus queletii (Schulzer) Vizzini, Simonini & Gelardi = Boletus queletii Schulzer ha pori arancio-rossastri, gambo giallastro, liscio o raramente decorato da piccole granulazioni bruno-rossastre nella parte mediana, color barbabietola alla base, carne virante al verde-bluastro e tipicamente barbabietola alla base del gambo. Note nomenclaturali Fungo largamente conosciuto con il binomio Boletus erythropus, creato da Persoon nel 1796 e sanzionato da Fries nel 1821, tuttavia non sarebbe corretto utilizzare tale epiteto, dal momento che solo nell'interpretazione di Fries corrisponde bene alla specie a cui lo associamo oggi, mentre nel concetto originale di Persoon era qualcosa di sicuramente diverso, la descrizione riconduce probabilmente ad un Suillellus queletii (il cappello viene definito su toni aranciati e il gambo è descritto come rosso alla base). Nell'anno 2014 indagini filogenetiche condotte dai micologi italiani Matteo Gelardi, Giampaolo Simonini e Alfredo Vizzini, hanno segregato la specie nel nuovo genere Neoboletus, genere le cui specie sono caratterizzate dalla carne che diventa blu al taglio, dai pori giallo-aranciati e dalla mancanza di reticolo sul gambo. Gli ultimi studi molecolari e tassonomici tendono ad attribuire i ritrovamenti di “Boletus erythropus” sotto conifere a Neoboletus praestigiator mentre i ritrovamenti sotto latifoglie sarebbero da attribuire secondo recenti indicazioni nomenclaturali a Neoboletus luridiformis. Quest’ultimo binomio nel corso delle prime revisioni era stato indicato quale prioritario, ma in base alle indicazioni tassonomiche formulate da alcuni autori, c'è chi sostiene che sia meglio accantonarlo e ritenerlo un sinonimo di Neoboletus xanthopus. Sono comunque tuttora in corso studi approfonditi sul genere Neoboletus i quali potrebbero portare ad ulteriori novità, ultimamente viene anche proposto come current name di Neoboletus luridiformis (Rostk.) Gelardi, Simonini & Vizziniun un nuovo taxa, Sutorius luridiformis (Rostk.) G. Wu & Zhu L. Yang 2016. Bibliografia DELLA MAGGIORA, M., 2016. Boletaceae Chevall. Stato attuale della nomenclatura , Atti XXXI Comitato Scientifico Regionale Associazione Gruppi Micologici Toscani, Annali Micologici. A.G.M.T. 9: 85-116. GANG, W., & BANG, F., & JIANPING, X., & XUE-TAI Z., & YAN-CHUN, L., & NIAN-KAI, Z., & MD. IQBAL, H., & ZHU, L. Y., 2014. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae Fungal Diversity DOI 10.1007/s13225-014-0283-8 [data di accesso 28/03/2017] INDEX FUNGORUM, VIZZINI, A.,2016. Index Fungorum no.300. [Data di accesso: 03/01/2022]. PERSOON, C.H., 1795. Observationes mycologicae. Annalen der Botanik (Usteri). 15:1-39. URBAN, A., & KLOFAC, W., 2015. Neoboletus xanthopus, a sibling species of Neoboletus luridiformis, and similar boletes with yellowish pileus colours. Sydowia 67: 175-187. Scheda di proprietà AMINT realizzata da Pietro Curti - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Lombardia; Maggio 2010; Foto di Massimo Biraghi.






-
Trametes ochracea (Pers.) Gilb. & Ryvarden 1987
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
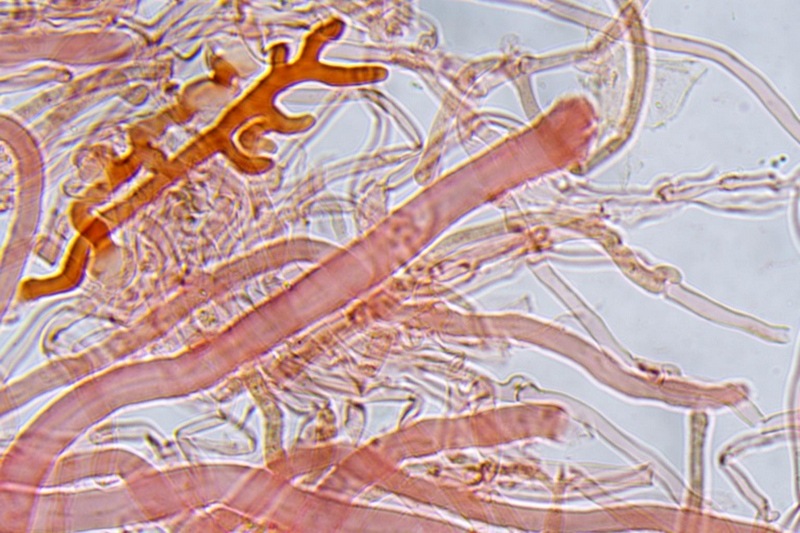
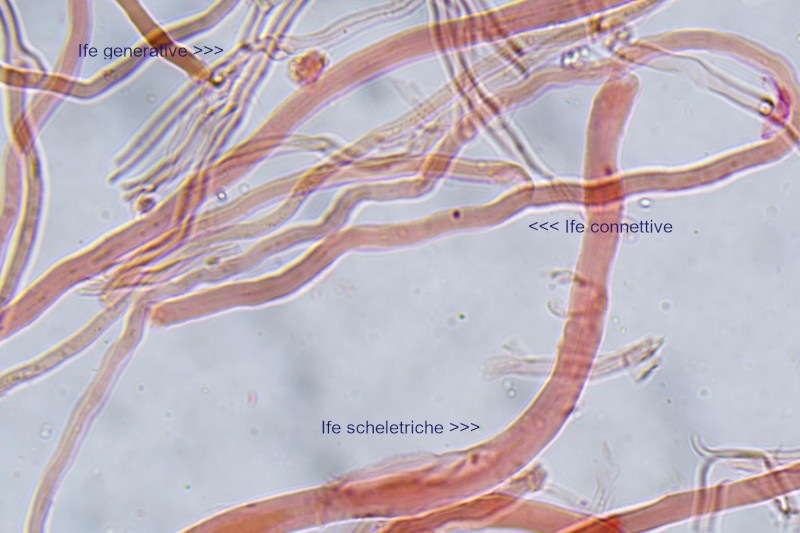
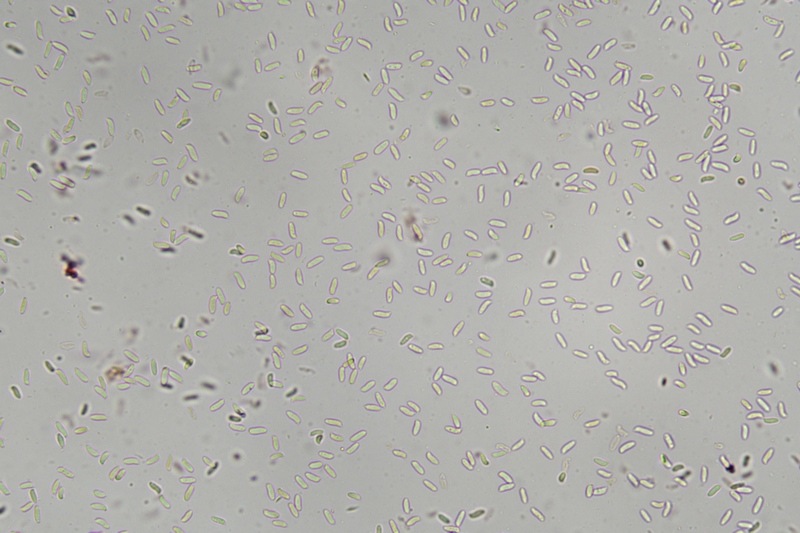
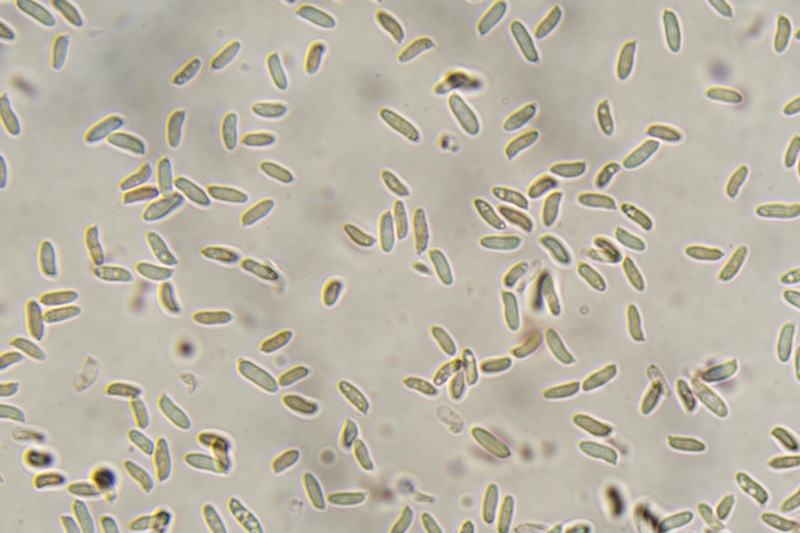
Trametes ochracea (Pers.) Gilb. & Ryvarden 1987 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Polyporales Famiglia Polyporaceae Sinonimi Polyporus zonatus (Nees : Fr.) Fr. 1821 Trametes multicolor (Schaeff.) Jülich 1982 Etimologia L'epiteto Trametes deriva dal latino Tràmes Tràmitis = passaggio, sentiero, percorso, viottolo, tramite; in riferimento alla conformazione dell'imenoforo che in alcune specie di questo genere si mostra vagamente a forma di labirinto. L'epiteto ochracea deriva dal latino ochra = ocra; per i cromatismi che contraddistinguono la specie. Descrizione I basidiomi, si sviluppano in colonie, sono sessili, a mensola, flabelliformi, dimidiati e spesso imbricati, con il margine intero, ripiegato sull’imenio con tonalità molto chiare. La superficie sterile è zonata, con sezioni tomentose alternate e altre tubercolate e a tratti vellutate, i colori comprendono vari cromatismi e sfumature, con tonalità che vanno dal marrone ocraceo a volte sbiadito, fino a zonature bruno nerastre. Il contesto è spesso, biancastro, sodo e senza alcuna linea nera che lo separi dal tomento; i tubuli sono corti e di colore ocraceo, i pori piccoli sono rotondi, ma si apprezzano anche pori più grandi e angolosi. Microscopia Sistema ifale trimitico formato da ife generatrici a pareti sottili, ialine, con evidenti GAF, larghe 1,5-2,2 µm, ife connettive tortuose, e ramificate a parete spessa di diamentro: 2,4-4,2 µm, e ife scheletriche a parete spessa, cilindriche o appena sinuose, con ingrossamenti ampolliformi, prive di setti, larghe 3-4,5 µm. Spore 5,6 [6,6; 6,9] 8,0 × 1,8 [2,3; 2,4] 2,9 µm; Q = 2,2 [2,8; 3,0] 3,6; N = 52; C = 95%; Media = 6,8 × 2,4 µm; Qm = 2,9; cilindriche o faseoliformi con apicolo prominente. Cuticola di tipo tricoderma con peli allungati. Habitat Cresce di norma su legno di latifoglie, mentre è più raro su quello di conifere. Si tratta di una specie ampiamente diffusa in Italia ed Europa, seppur meno frequente della più comune Trametes versicolor. Commestibilità e Tossicità Non commestibile. Specie simili Trametes versicolor (L. : Fr.) Lloyd si distingue per il basidioma meno ispessito e l'attaccatura al substrato più sottile, per le zonature più vistose, tendenti al nerastro o bluastro e per la presenza di una linea nera tra contesto e tomento. Trametes hirsuta (Wulfen : Fr.) Lloyd si distingue per il contesto più sottile e per la superficie sterile non zonata o solo leggermente zonata a bande concolori cremeo-biancastre. Bibliografia BREITENBACH, J. & KRANZLIN, F., 1986. Champignons de Suisse. Vol. 2. Champignos sans lames. Lucerna. Ed. Verlag Mykologia. BERNICCHIA, A., 2005. Polyporaceae s.l. Fungi Europæi. Vol 10. Alassio (SV): Ed. Candusso. BERNICCHIA, A. & GORJÓN, S.P., 2020. POLYPORES of the Mediterranean Region. ROMAR. Scheda di proprietà AMINT realizzata da Giovanni Galeotti e Stefano Rocchi. Approvata e revisionata dal CLR Micologico di AMINT. Regione Umbria; Città di Castello (PG); Gennaio 2021; Foto e microscopia di Giovanni Galeotti. Esemplari rivenuti su tronco di latifoglia in colonie presenti anche sui rami apicali. (Exsiccatum GG210201-01) ) In evidenza l'imenoforo con pori piccoli rotondeggianti, a tratti angolosi e margine sottile biancastro leggermente involuto. Sezione dell'imenoforo. Si evidenzia l'assenza della linea nerastra, tra la carne e i tubuli. Sistema ifale trimitico. Osservazione in Rosso Congo, a 400×. Ife connettive tortuose e ramificate a parete spessa. Osservazione in Rosso Congo, a 400×. Sistema ifale. Osservazione in Rosso Congo, a 1000×. Giunti a fibbia presenti sulle ife generatrici. Osservazione in Rosso Congo, a 1000×. Si evidenziano graficamente le ife: Ife generative; Ife connettive; Ife scheletriche. Spore. Osservazione in acqua, a 400×. Spore 5,6 [6,6; 6,9] 8,0 × 1,8 [2,3; 2,4] 2,9 µm; Q = 2,2 [2,8; 3,0] 3,6; N = 52; C = 95%; Media = 6,8 × 2,4 µm; Qm = 2,9. Osservazione in acqua, a 1000×. Basidi e basidioli. Osservazione in Rosso Congo, a 1000×. Giunti a fibbia presenti alla base dei basidi e basidioli. Osservazione in Rosso Congo, a 1000×. Cuticola. Osservazione in Rosso Congo, a 100×. Cuticola. Osservazione in Rosso Congo, a 400×.




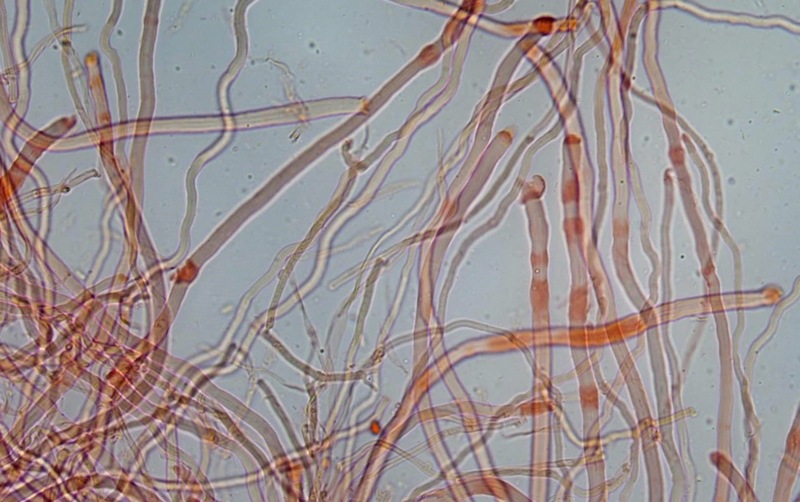
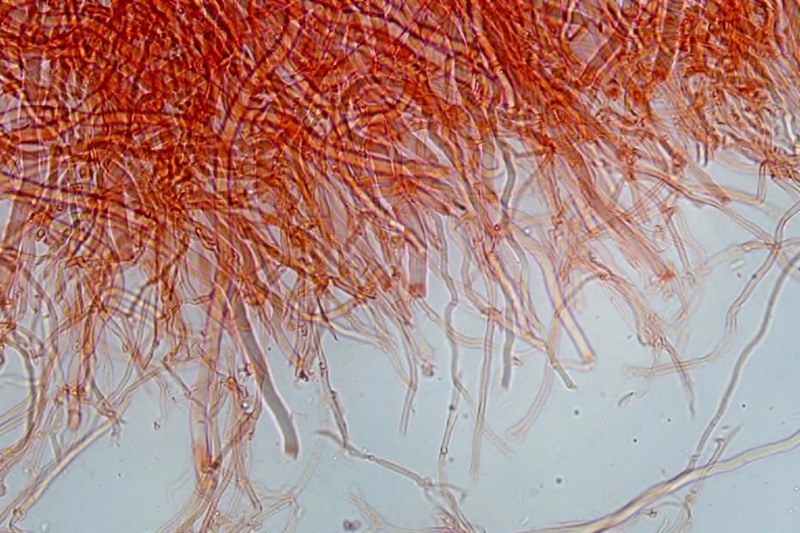
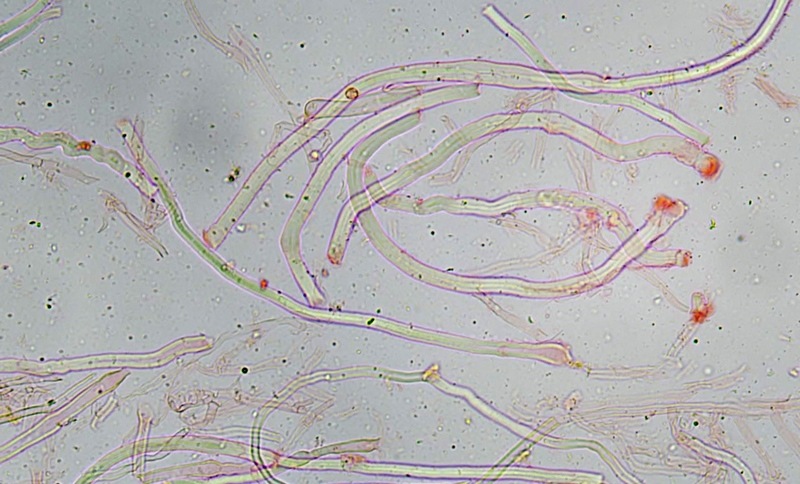
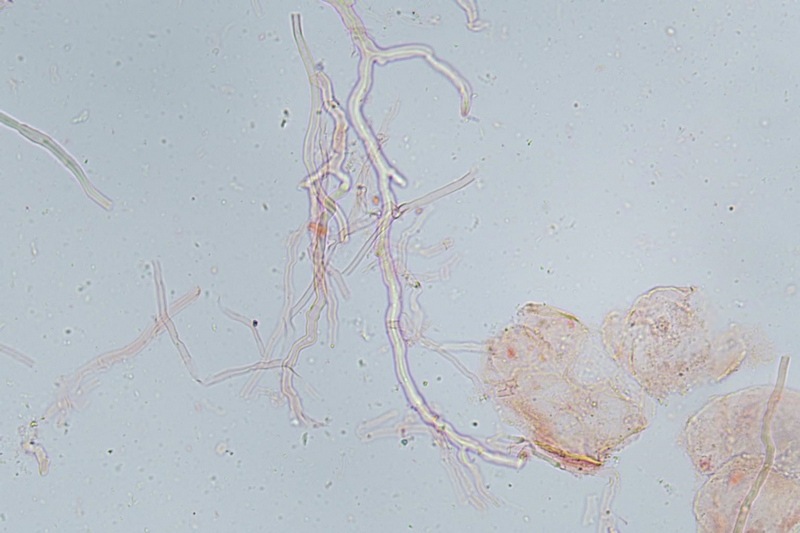
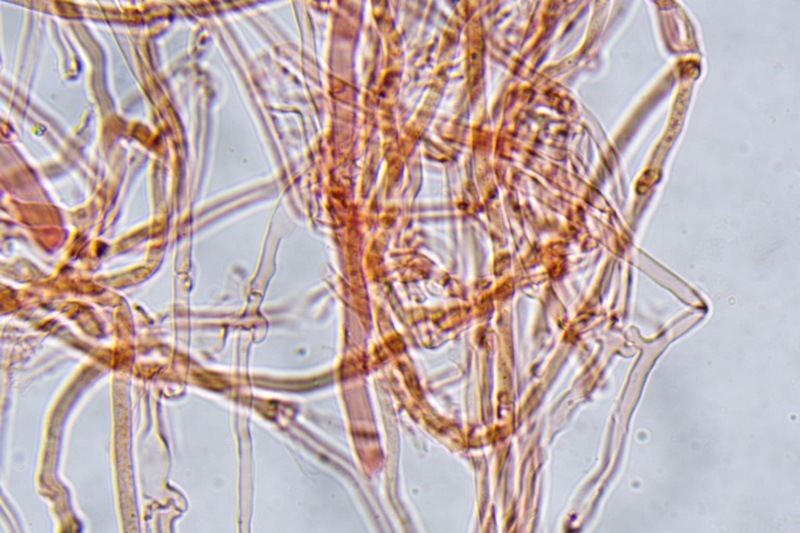
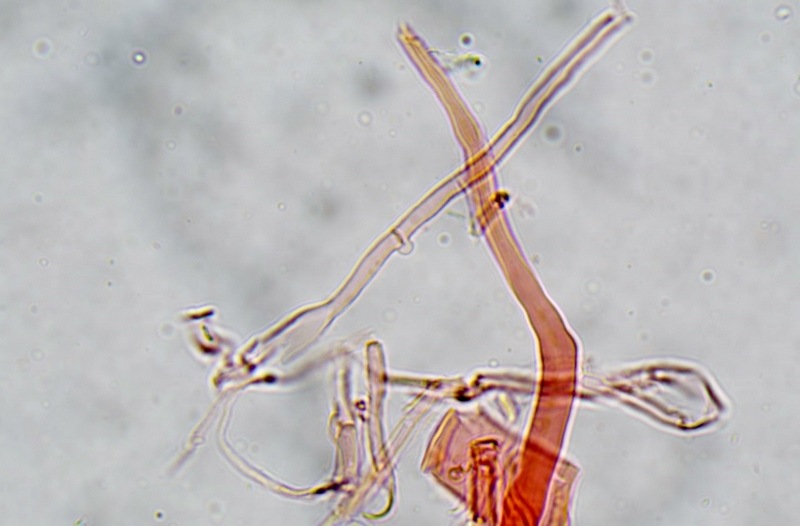
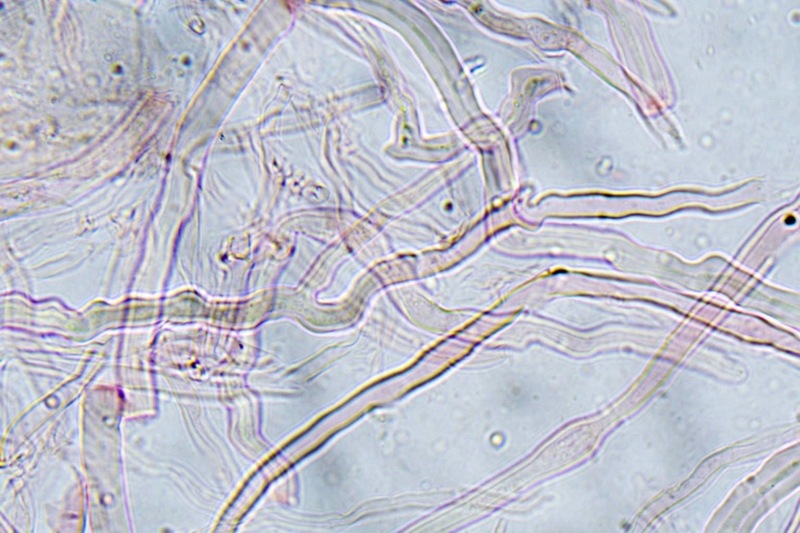
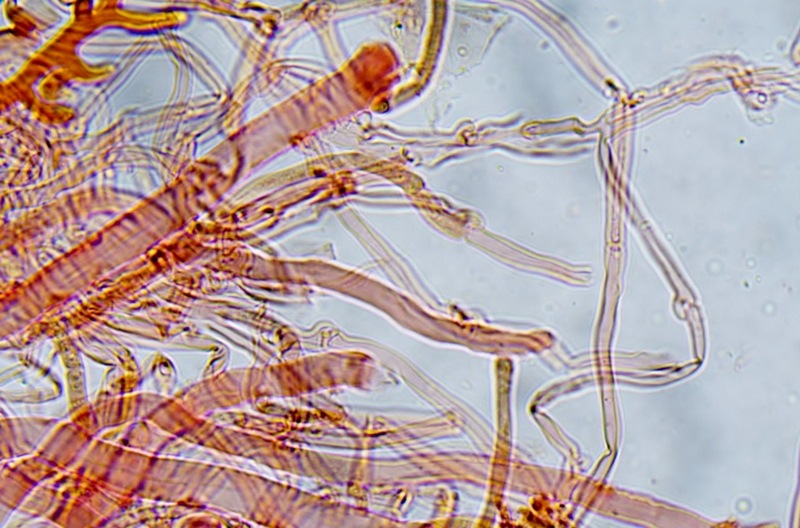
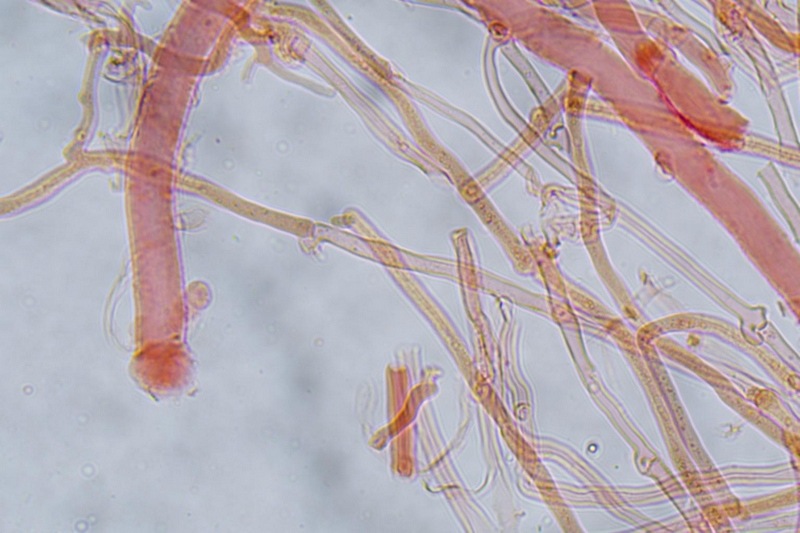
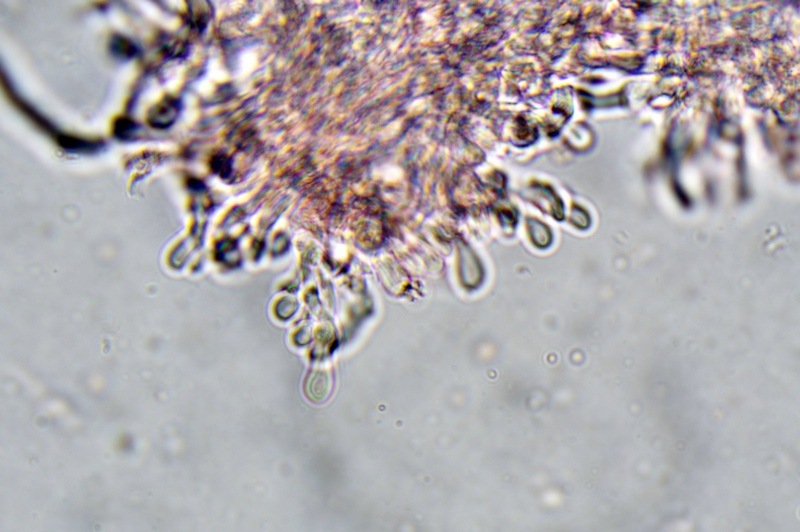
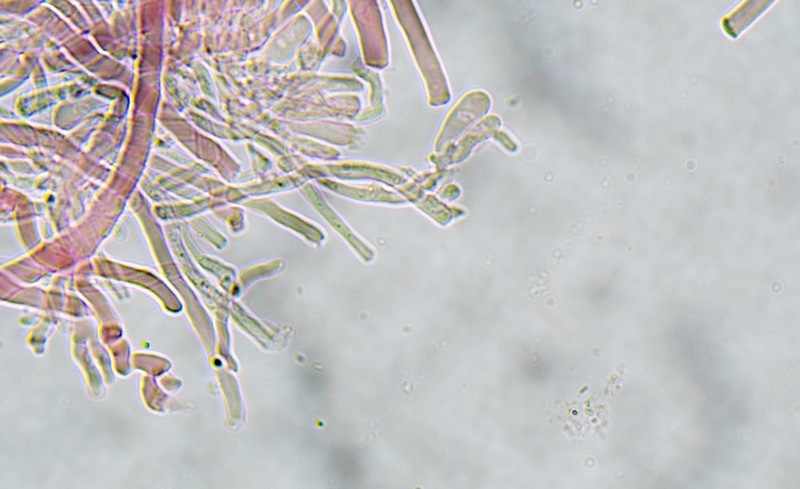
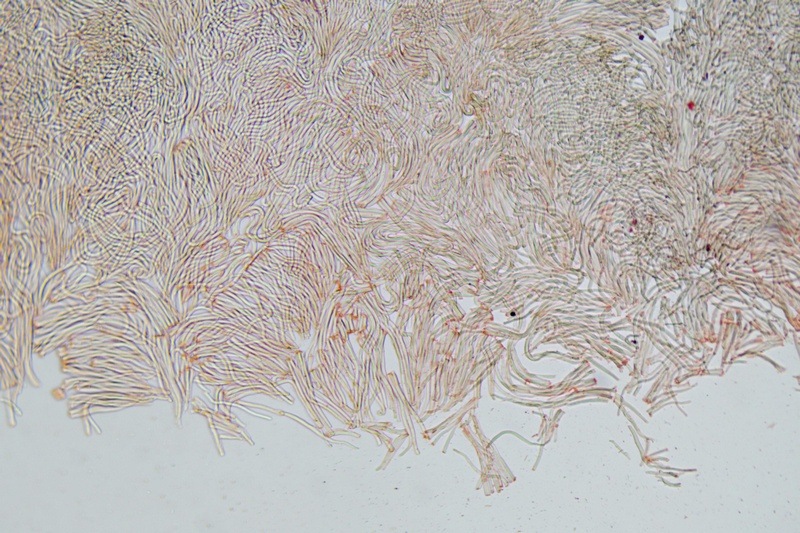

-
Amylosporus campbellii (Berk.) Ryvarden 1977
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Amylosporus campbellii (Berk.) Ryvarden 1977 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Russulales Famiglia Bondarzewiaceae Sinonimi Polyporus campbellii Berk. 1854 Polyporus anthelminticus Berk. 1866 Polyporus popanoides Cooke 1881 Tyromyces graminicola Murrill 1915 Polyporus graminicola (Murrill) Murrill 1915 Polyporus mollitextus Lloyd 1919 Polyporus propinquus Lloyd 1922 Polyporus hypomiltinus Bres. 1937 Scutiger tisdalei Murrill 1943 Polyporus tisdalei (Murrill) Murrill 1943 Amylosporus graminicola (Murrill) Ryvarden 1973 Wrightoporia campbellii (Berk.) Teixeira 1992 Etimologia L'epiteto Amylosporus deriva dal greco ἄμυλος [ámylos] = amido, e σπορά [sporá] = seme, per le spore amiloidi. L'epiteto campbellii è dedicato al micologo Campbell. Descrizione Basidiocarpi a crescita annuale, si comportano da parassiti e presumibilmente anche da saprofiti, carnosi ed acquosi da freschi, fragili, spugnosi e leggeri ad essiccazione, sessili o muniti di gambo breve, tozzo, appena abbozzato; a crescita lignicola o apparentemente terricola. Si presentano in forma singola, sporgenti, a forma di mensola o circolare, o sovrapposti l’uno all’altro. In questo caso, i nuovi esemplari crescono su quelli già esistenti in fase di degradazione, oppure confluenti fino a formare un agglomerato di dimensioni consistenti. Superficie sterile liscia, lucida, asciutta, con leggeri solchi che delineano piccole aree irregolari inizialmente di colore bianco o biancastro, che tende poi, verso la maturazione, ad assumere una colorazione rosato-ocracea più intensa, immutabile al tocco. Margine ottuso, ondulato, a volte lobato con lobi poco accentuati, tanto da assumere una conformazione subglobosa ed irregolare, di colore rosa chiaro, come la superficie, negli esemplari giovani, ocracea più o meno intensa in quelli più maturi. Superficie fertile a tubuli monostratificati di media lunghezza con pori piccoli, rotondi o leggermente allungati in maniera irregolare, colore bianco virante al rosa al tocco, tendente a scurire verso la maturazione. Gambo assente o, se presente, tozzo, corto, appena abbozzato, centrale o laterale. Carne morbida ed elastica, fibrosa, omogenea, bianca, acquosa e carnosa nel fresco; spugnosa, leggera e fragile dopo essiccazione. Odore gradevole, con intensità più accentuata negli esemplari giovani; sapore dolce, fungino, di nocciola. Microscopia Sistema ifale dimitico caratterizzato da ife generatrici ialine, doppie e verticillate, a parete sottile, vacuolate con setti e unioni a fibbia semplici, e da ife scheletriche, anch’esse ialine a parete spessa, diritte o con andamento più o meno ondulato, moderatamente ramificate, non destrinoidi al reagente di Melzer. Spore (4,4) 4,5-5,5 (6,3) × (3,5) 3,7-4,3 (4,8) µm; Meda 5 × 4 µm; Q = (1,1) 1,14-1,4 (1,5); Qm = 1,3; amiloidi, con forma ellissoidale, a parete spessa e finemente echinulate. Quest'ultima caratteristica non è facile da osservare, tant'è che sembrano lisce. Habitat Specie tipicamente diffusa nelle aree tropicali e subtropicali, mai associata a colture arboree o arbustive boschive. Fruttifica, stante la nostra recente esperienza ed a quanto riportato in letteratura, nelle intercapedini della pavimentazione, attaccato ai muretti di delimitazione delle aiuole, nei giardini, nei parchi, nei viali alberati, in associazione a piante ornamentali, come Chamaerops humilis L. (Palma nana), Thuja orientalis (L.) Franco, e anche su ceppo di Pino domestico (Pinus pinea L.) e su Palma messicana [Washingtonia robusta (Lindl.) H. Wendl.]; inoltre, collegato presumibilmente ai residui radicali di pianta ornamentale di Pittosporum s.l., per mezzo di un gambo rudimentale, posizionato lateralmente al basidioma, oppure inglobante ramoscelli di rosmarino (Rosmarinus officinalis Schleid.). Commestibilità e Tossicità Non commestibile. Genere Amylosporus Nel genere, la cui specie tipo è Amylosporus campbellii, trovano posto altri 10 taxa: Amylosporus auxiliadorae Drechsler-Santos & Ryvarden; Amylosporus bracei (Murrill) A. David & Rajchenb.; Amylosporus casuarinicola (Y.C. Dai & B.K. Cui) Y.C. Dai, Jia J. Chen & B.K. Cui; Amylosporus efibulatus (I. Lindblad & Ryvarden) Y.C. Dai, Jia Chen J. & B.K. Cui; Amylosporus guaraniticus Campi & Robledo; Amylosporus rubellus (Y.C. Dai) Y.C. Dai, Jia Chen J. & B.K. Cui; Amylosporus ryvardenii Stalpers; Amylosporus succulentus Jia J. Chen e L.L. Shen; (Campi et al., 2017; IF, 2020; MB, 2020); Amylosporus daedaliformis G.Y. Zheng & Z.S .Bi; Amylosporus wrihtii Rajchenb. (IF, 2020; MB, 2020). Tali specie sono tutte originarie di aree tropicali e sub-tropicali, a tipica crescita lignicola. Presentano cappello di colore bianco-biancastro, lucido e tendente, verso la maturazione, ad assumere colorazione ocracea. Generalmente sono sessili o hanno gambo breve, grossolano, tozzo, appena abbozzato. La zona imeniale, a tubuli, è caratterizzata da pori piccoli, circolari o irregolarmente allungati, di colore bianco-biancastro. Il sistema ifale, dimitico, è caratterizzato dalla presenza di ife generatrici ialine e ife scheletriche. I basidi tetrasporici hanno conformazione clavata; le spore, di forma ellissoidale, sono lisce o leggermente verrucose, amiloidi. Osservazioni I numerosi e recenti ritrovamenti lasciano facilmente dedurre che la specie si stia facilmente acclimatando alle nostre latitudini, ed in maniera abbondante; fattore, questo, di notevole importanza che si pone quale indicatore dell’attuale processo di mutazione climatica che, ormai da diversi anni, caratterizza le regioni del mediterraneo, sempre più vicine alle caratteristiche climatiche delle zone tropicali e sub-tropicali. Bibliografia BERNICCHIA, A., GENOVESE, R. & GORJÓN S.P., 2017. Amylosporus campbellii (Berk.) Ryvarden (Bondarzewiaceae, Basidiomycota), new record to Europe. Mycosphere, Journal of Fungal Biology 8 (1): 98–101. BERNICCHIA, A. & GORJÓN S.P., 2020: Polypores of the Mediterranean Region. Romar Edizioni, Segrate Milano, I. CAMPI, M., MAUBET, Y., GRASSI, E. & ROBLEDO, G., 2017. Amylosporus guaraniticus sp. nov. (Wrightoporiaceae, Russulales) a new neotropical species in Paraguay. Mycosphere 8 (6): 1060–1070. Doi 10.5943/mycosphere/8/6/6. CHEN, J.J. & SHEN, L.L., 2014. Amylosporus succulentus sp. nov. (Russulales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Cryptogamie, Mycologie 35 (3): 271–282. INDEX FUNGORUM, 2021. Amylosporus campbellii. [Data di accesso: 15/04/2021]. MICELI, A. & DI VINCENZO, C., 2019. Amylosporus campbellii (Berk.) Ryvarden (1977). Primo ritrovamento nel territorio messinese. RMR, Boll. AMER 111 XXXVI (3): 135–141. MYCOBANK, 2021. Amylosporus campbellii. [Data di accesso: 15/04/2021]. VERMA, R.K., TIWARI, C.K., PARIHAR, J. & KUMAR, S., 2016. Diversity of Amylosporus campbellii in central India. Van Sangyan 3 (11) : 20–23. Tropical Forest Research Institute, Jabalpur, MP, India. Scheda realizzata da Angelo Miceli, Carmelo Di Vincenzo e Tomaso Lezzi - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Sicilia, Messina, comune di Rometta, loc. Rometta Marea; Settembre 2020; Foto di Angelo Miceli e Carmelo Di Vincenzo. La raccolta seguente presentava una larghezza da 8 a 18 cm e una lunghezza da 6 a 17 cm. Secondo Bernicchia et al. (2017, 2020) raggiungono anche 30-50 cm di larghezza). La maturazione completa viene raggiunta in 30-50 giorni circa. Ritrovamento avvenuto a circa 20 m s.l.m. e a circa 250 m di distanza dalla spiaggia, in due stazioni di crescita posizionate a 200 m di distanza tra loro, una nel Complesso “Il Gabbiano”, l’altra nel Complesso “Due Torri”, in spazi esterni di fabbricati a civile abitazione caratterizzati dalla presenza di piante ornamentali di diversa natura. Nella prima stazione, il 21.7.2020, due esemplari, sporgenti a forma di mensola dal muretto di recinzione di una aiuola, legati, presumibilmente, ai residui radicali di pianta ornamentale di Pittosporum s.l. (Pittosporo), creando il collegamento per mezzo di un gambo rudimentale, posizionato lateralmente al basidioma, leg. A. Miceli e C. Di Vincenzo (Centro di Cultura Micologica, Messina). Nella seconda stazione, il 12.8.2020, numerosi esemplari distribuiti in un giardino con piante ornamentali appartenenti a generi e specie diverse, cresciuti singolarmente o raggruppati in diversi esemplari, posizionati nelle aiuole, nelle intercapedini della pavimentazione, attaccati al muretto di delimitazione delle aiuole, inglobanti ramoscelli di rosmarino (Rosmarinus officinalis) o, più probabilmente, collegati alle radici di un ceppo residuale di Pinus pinea, leg. A. Miceli e C. Di Vincenzo. Amylosporus campbellii. Esemplari variamente concresciuti. Foto di Angelo Miceli. Particolare della superficie fertile. Foto di Carmelo Di Vincenzo. Sistema ifale. Osservazione in Rosso Congo, a 400×. Foto di Carmelo Di Vincenzo. Ife generative. Osservazione in Rosso Congo, a 400×. Foto di Carmelo Di Vincenzo. Ifa scheletrica. Osservazione in Rosso Congo, a 400×. Foto di Carmelo Di Vincenzo. Spore, Amiloidia. Osservazione in Reattivo di Melzer, a 400×. Foto di Carmelo Di Vincenzo.








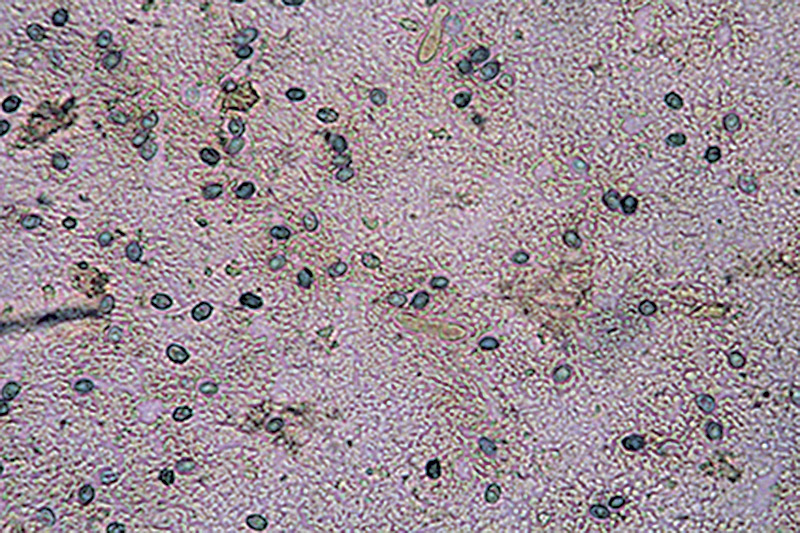
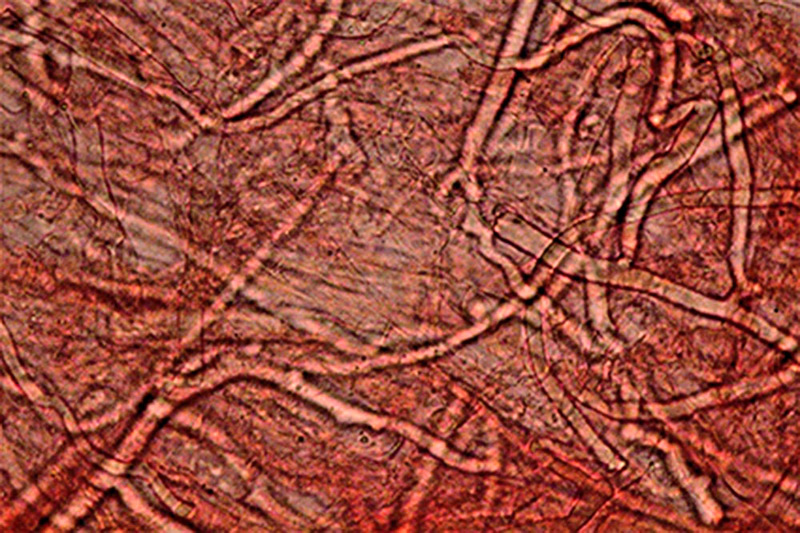

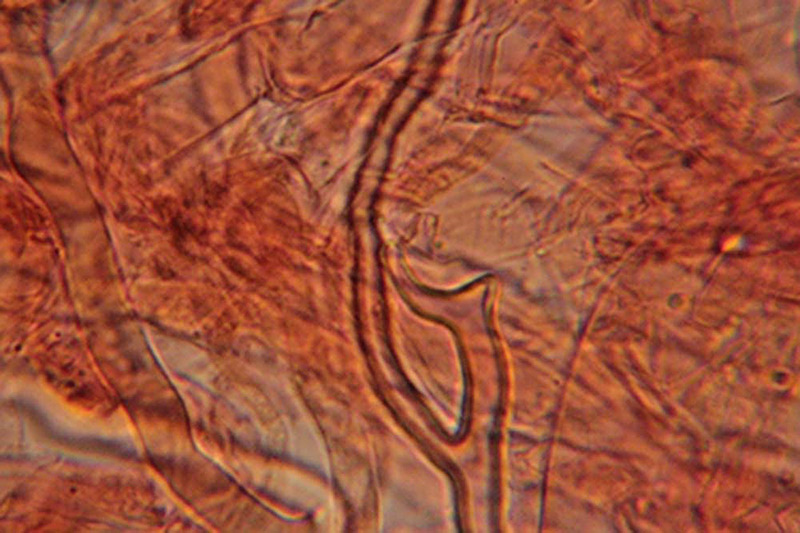
-
Amanita porrinensis Freire & M.L. Castro ex M.L. Castro 1998
Archivio Micologico ha postato un topic nell'area Funghi Velenosi
Amanita porrinensis Freire & M.L. Castro ex M.L. Castro 1998 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Amanitaceae Genere Amanita Sottogenere Amanitina Sezione Phalloideae Sinonimi Amanita porrinensis Freire & M.L. Castro 1987 Note tassonomiche Il primo ritrovamento di questa specie è datato 10 ottobre 1984, ed è stato effettuato nel territorio boschivo del comune di Vigo, in località Madroa, nella provincia di Pontevedra, in Spagna, ai margini di un’area boschiva a coltura mista. Successivamente, nel 1987, i micologi Luis Freire e Maria Luisa Castro ne danno notizia presentandola come “specie nuova” in Anales Jardìn Botànico de Madrid (1987), provvedendo ad una prima descrizione della specie alla quale viene attribuito il binomio Amanita porrinensis Freire & M.L. Castro. Nel contesto viene precisato che “la possibilità che si tratti di un nuovo taxon del genere Amanita è stata confermata dai dottori Moreno e Bon; tuttavia, si rimane in attesa della conferma del dottor Bas, specialista in questo genere” (Freire & Castro 1987]. Tuttavia, la specie viene presentata e descritta in maniera incompleta, priva di numero identificativo e della menzione dell’erbario ove l’holotypus è stato depositato, e quindi non ritenuta valida (nomen invalidum) ai fini della conferma del taxon (Neville & Poumarat, 2004; IF, 2018; MB, 2018]. Per questo motivo, il taxon viene riproposto successivamente da Castro (1998), mediante pubblicazione rispondente ai requisiti richiesti dal Codice di Nomenclatura, su Mykes, Bollettino del Gruppo Micologico Galego, e definitivamente accettato dal mondo scientifico internazionale come Amanita porrinensis Freire & M.L. Castro ex M.L. Castro. Etimologia L'epiteto Amanita deriva dal greco Ἀμανὶτης [Amanitos] = fungo del monte Amano, da Ἄμανος [Amanos] = Amano, catena di monti tra la Cilicia e la Siria, nella Turchia asiatica, dove questo genere sembra fosse abbondante in antichità. L'epiteto porrinensis deriva dalla località Porriño (Presso Vigo, Spagna) e dall'estensione latina -ensis = proveniente da Porriño. Cappello Di piccole dimensioni, 2-8 cm, inizialmente semisferico, campanulato verso la maturazione, spianato-convesso, caratterizzato da un largo umbone centrale molto prominente che gli conferisce spesso un tipico aspetto a “sombrero messicano”; margine sottile, inizialmente liscio poi, verso la maturazione, leggermente striato e incurvato, privo di residui di velo primario (verruche). Cuticola liscia, leggermente viscida a tempo umido, bianca, immutabile, difficilmente separabile. Imenoforo Costituito da lamelle bianche con riflessi rosati, fitte, sublibere al gambo, intervallate da lamellule. Spore in massa bianche. Gambo Bianco, centrale, cilindrico clavato, anche obeso, con bulbo basale subgloboso, con bande sericee trasversali, ricoperto da fiocchetti bianchi macchiati di ocra, fistoloso per tutta la sua lunghezza. Anello Fragile, membranoso e sottile, visibile negli esemplari giovani, poco evidente in quelli maturi dove si presenta sotto forma di lembi aderenti al gambo e sulle lamelle in maniera poco visibile. Volva Bianca, sottile ed aderente al gambo, gracile e poco persistente, si disgrega facilmente a maturazione avanzata ed al tocco. Carne Bianca, sottile nel cappello, spessa nel gambo e di consistenza spugnosa, leggermente imbrunente, alla sezione, verso la base del gambo, inodore. Reazione macrochimiche Reazione gialla sulla cuticola con basi forti (KOH e NaOH). Habitat Tipicamente autunnale. Pino domestico (Pinus pinea), Pino marittimo (Pinus pinaster), Pino d’Aleppo (Pinus halepensis) Castagno (Castanea sativa), Leccio (Quercus ilex), Roverella (Quercus pubescens), con presenza di macchia mediterranea con predominanza di Erica, Cisto, Corbezzolo e Ginestra. Commestibilità e tossicità Specie velenosa mortale, come le altre della sezione Phalloideae: Amanita phalloides (Vaill. ex Fr. : Fr.) Link, Amanita phalloides var. alba Costantin & L.M. Dufour, Amanita verna Bull. ex Lam. : Fr. ed Amanita virosa Bertill., con le quali va a completare, per il contenuto degli stessi principi tossici, il quadro delle Amanita mortali. È causa di sindrome falloidea caratterizzata soprattutto da grave insufficienza epatica. Specie simili Amanita porrinensis, soprattutto per la colorazione generale bianco-biancastra, presenta una certa affinità con altre specie velenoso-mortali appartenenti alla sezione Phalloideae del sottogene Amanitina, se ne distingue morfologicamente per la conformazione del cappello spesso a “sombrero messicano” e per altre caratteristiche minori. Amanita phalloides var. alba differisce per la conformazione del cappello piano-convesso e mai umbonato; per il gambo zebrato, mai ricoperto da squamule; per l’anello sempre evidente e persistente, mai fugace; per il bulbo basale grosso e prominente; per la volva sempre evidente, membranosa e persistente, aderente al bulbo e libera all’apice. Amanita verna differisce per la conformazione del cappello piano-convesso e mai umbonato; per il gambo liscio, mai ricoperto da squamule; per il bulbo basale grosso e prominente; per la volva interamente avvolgente il bulbo e libera all’apice. Amanita virosa è la specie che presenta maggiori affinità con Amanita porrinensis, soprattutto per la conformazione del cappello che, inizialmente, è ovoidale-campanulato e, poi, campanulato convesso, a volte con umbone ottuso a maturazione, ma mai con la tipica conformazione spesso a “sombrero messicano”; per la struttura dell’anello poco persistente e fugace che, come per Amanita porrinensis, lascia residui sul margine del cappello e sul filo lamellare; per le fioccosità presenti sul gambo che, anche se più abbondanti, la rendono molto vicina ad Amanita porrinensis. Differisce per il colore del cappello che, come per Amanita porrinensis, è bianco-biancastro, ma con sfumature ocra-rosate nella zona discale specialmente negli esemplari maturi; per il bulbo basale prominente e globoso; per la volva interamente avvolgente il bulbo, ampia e membranosa, poco libera al gambo ma persistente e sempre presente anche a maturazione avanzata. Osservazioni Della stessa specie sono noti al momento pochi ritrovamenti, tra cui: 10 ottobre 1984, ritrovamento dell’holotypus, in località Madroa, comune di Vigo, Provincia di Pontevedra (Spagna), al margine di bosco misto (Quercus robur e Pinus pinaster), da Jaime Diz (gruppo M. Porriño) (Freire & Castro, 1987; Neville et al., 2000). 11 novembre 1991, in località Picouto, sempre nel comune di Vigo, Provincia di Pontevedra (Spagna), in bosco misto da Jaime Diz (Castro & Blanco-Dios, 2007). 16 ottobre 1998, in località Monte Cistorello, Colli Berici, Comune di Sossano (VI), in bosco misto di querce, carpini e castagni. Ritrovamento effettuato da Franco Gasparini e Franco Serafin (Gasparini & Serafin, 2012). 17 ottobre 1998, in località Volano, comune di Comacchio (FE) a 0 metri s.l.m., in bosco misto formato da Pinus pinea, Pinus pinaster con limitata presenza di Quercus ilex. Ritrovamento effettuato da Gianni Monterumici (Neville et al., 2000). Ulteriori ritrovamenti, senza precisazione temporale, sono stati segnali nel Canton Ticino (Svizzera) e, in Italia sui Colli Euganei (Galli, 2007). Bibliografia CASTRO, M.L., 1998. Amanita porrinensis L. Freire et M.L. Castro, estudio comparativo con outros taxons da sección Phalloideae (Fr.) Quél. Mykes. 1: 57-60. FREIRE, L. & CASTRO, M.L., 1987. Nueva especie del género "Amanita". Anales del Jardín Botánico de Madrid. 44(2): 533-534. GALLI, R., 2007. Le Amanite. 2a edizione. Ed. Dalla Natura, Milano. GASPARINI, F. & SERAFIN, F., 2012. La quarta Amanita mortale. Bollettino del Gruppo AMB di Vicenza. XXXII 2: 40-42. INDEX FUNGORUM, 2021. Amanita porrinensis. [Data di accesso: 06/04/2021]. MICELI, A., 2019. Amanita porrinensis, una specie rarissima ritrovata sui Monti Peloritani. MicoPonte (Bollettino del Gruppo Micologico Massimiliano Danesi) 12: 27-33. [Data di accesso: 06/04/2021]. MYCOBANK, 2021. Amanita porrinensis. [Data di accesso: 06/04/2021]. NEVILLE, P. & POUMARAT, S., 2004. Amanitae. Amanita, Limacella & Torrendia. Fungi Europaei 9. Ed. Candusso, Alassio (SV). NEVILLE, P.m POUMARAT, S. & MONTERUMICI, G., 2000: Una rara Amanita della sezione Phalloideae, nuova per l’Italia: Amanita porrinensis. Bollettino del Gruppo Micologico G. Bresadola – Nuova Serie. 43 (2): 143-150. Trento. Scheda realizzata da Angelo Miceli e Tomaso Lezzi - Approvata e Revisionata dal CLR Micologico di AMINT. Regione Sicilia, Monti Peloritani; Ottobre 2018; Foto di Angelo Miceli e Franco Mondello e descrizione di Angelo Miceli. In data 27 ottobre 2018 ad opera di Mario Mondello e Piero Battaglia, soci del Centro di Cultura Micologica di Messina, nel comune di Rometta (ME), Monti Peloritani sulla dorsale Dinnammare-Monte Scuderi in prossimità del sentiero Girasì, sul versante ovest del monte Pizzo Bandiera a circa 950 m s.l.m. vengono trovati tre esemplari, cresciuti in un raggio di circa 30 metri. Le dimensioni sono circa 8 cm di altezza, cappello 2-4 cm, in un bosco misto formato da Pinus pinea, Pinus pinaster, Quercus ilex con sottobosco di Erica arborea. Si segnala un ulteriore ritrovamento, nella stessa stazione di crescita, di un unico esemplare, in data 21 novembre 2018 ad opera del micologo Franco Mondello, di Angelo Miceli e Mario Mondello, soci dello stesso Centro di cultura Micologica di Messina. Habitat di ritrovamento: Pino domestico (Pinus pinea), Pino marittimo (Pinus pinaster), Pino d’Aleppo (Pinus halepensis) Castagno (Castanea sativa), Leccio (Quercus ilex), Roverella (Quercus pubescens), con presenza di macchia mediterranea con predominanza di Erica, Cisto, Corbezzolo e Ginestra. Foto di Angelo Miceli. Foto di Franco Mondello. Foto di Angelo Miceli. Vista d'insieme del carpoforo. Cappello. Anello che si sta distaccando dalle lamelle. Gambo fioccoso. Bulbo basale e volva.











-
Coprinellus congregatus (Bull.) P. Karst. 1879
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
Coprinellus congregatus (Bull.) P. Karst.; Regione Lombardia; Marzo 2021; Foto macro- e micro- scopiche di Sergio Mombrini. Secondo ritrovamento alcuni giorni dopo il precedente, in altra località, sempre su letamaio, microscopia come il ritrovamento precedente. -
Coprinellus congregatus (Bull.) P. Karst. 1879
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Coprinellus congregatus (Bull.) P. Karst. 1879 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Psathyrellaceae Sinonimi Coprinus congregatus (Bull.) Fr. 1838 Coprinus congregatus var. apleurocystidiosus Bogart 1975 Coprinus congregatus var. megasporus Bogart 1975 Tulosesus congregatus (Bulliard) D. Wächt. & A. Melzer 2020 Etimologia L'epiteto Coprinellus deriva dal nome di genere Coprinus e dal suffisso diminutivo latino -ellus, per l'apetto simile a piccoli Coprinus. L'epiteto congregatus deriva dal latino congregatus = riunito, aggregato, per la crescita in più esemplari riuniti. Cappello Chiuso fino a 16 × 12 mm, espanso con diametro fino a circa 28 mm, pubescente, velo assente. Lamelle Libere al gambo, presto grigie e infine nere e deliquescenti. Gambo Fino a 50-60 × 1,5-3 mm, pubescente, bianco. Carne Esigua, odore assente. Habitat Fungo coprofilo. Microscopia Spore 11,8-14,1 × 6,3-7,2 µm; Qm=1,9; ellissoidali-ovoidali; poro germinativo eccentrico. Basidi 21-32 × 8,5-11 µm; tetrasporici. Pseudoparafisi 5-6 per basidio. Cheilocistidi 30-80 × 20-70; globosi, ellissoidali. Pleurocistidi 90-110 × 50-60 µm; ellissoidali, leggermente utriformi. Pileipellis un imeniderma composto da elementi ellissoidali, arrotondati. Pileocistidi (setole-setae) 60-120 × 11-17 × 3,5-7,5 µm; lageniformi, attenuati. Caulocistidi 53-115 × 6-17 × 3-5 µm, lageniformi, attenuati. Sclerocistidi assenti. GAF assenti. Commestibilità e tossicità Non commestibile. Osservazioni Per quanto concerne la nomenclatura e alcune delle sinonimie si è fatto riferimento a Funga Nordica (2012), IF utilizza anch'esso il binomio Coprinellus congregatus e lo considera ancora prioritario, anche se inserisce anche Tulosesus congregatus con lo stesso basionimo Agaricus congregatus, ma stranamente senza mettere in relazione Coprinellus congregatus con Tulosesus congregatus, probabilmente al momento (1 Aprile 2021) i due taxa non sono ancora ben allineati; mentre Mycobank tenendo conto del recente lavoro di Dieter Wächter & Andreas Melzer (2020) aggiorna la tassonomia a Tulosesus congregatus (Bulliard) D. Wächt. & A. Melzer. Specie simili Coprinellus ephemerus (Bull.) Redhead, Vilgalys & Moncalvo è un sosia perfetto, per quanto riguarda le differenze tra i due citiamo le osservazioni ben riassunte e pubblicate in Caccialli et al. (1999): "Secondo Citerin (1992) la crescita cespitosa, la mancanza di giunti a fibbia e di cheilocistidi lageniformi, simili alle setole pileiche, sono in grado di inquadrare Coprinellus congregatus e di distinguerlo da Coprinellus ephemerus. Secondo altri studiosi (Moreno & Faus, 1984; Uljé & Bas, 1991) l'unico criterio distintivo sicuramente valido va ricercato nella presenza o meno di giunti a fibbia, soprattutto a livello miceliare". Per quanto concerne lo studio dei campioni in esame non sono stati trovati GAF nelle ife miceliari, mentre erano presenti invece numerosi falsi GAF rinvenuti soprattutto nelle ife verso la base del gambo. Bibliografia AA.VV., 2012. Funga Nordica. Agaricoid, boletoid and cyphelloid genera. Ed. Nordsvamp. CACIALLI, G., CAROTI, V. & DOVERI, F., 1999. Contributio ad Cognitionem Coprinorum. Monografie di Pagine di Micologia 1: 1–256. CACIALLI, G., CAROTI, V. & DOVERI, F., 2002. Aggiornamento della Monografia "Contributio ad Cognitionem Coprinorum". Mycologia Montenegrina V, Separatum: 119–129. CITÉRIN, M. 1992. Clé analytique du genre Coprinus Pers. Doc. Mycol. 22(86): 1–28. KARSTEN, P.A., 1879. Rysslands, Finlands och den Skandinaviska halföns Hattsvampar. Förra Delen: Skifsvampar. Bidrag till Kännedom av Finlands Natur och Folk. 32: 1-571. MORENO, G. & FAUS, J., 1984. Tres especies raras del genero Coprinus (Agaricales) de Cataluña, España. Cryptogamie, Mycologie 5: 3–17. REDHEAD, S.A., VILGALYS, R., MONCALVO, J.-M., JOHNSON, J. & HOPPLE, J.S. JR., 2001. Coprinus Persoon and the disposition of Coprinus species sensu lato. Taxon. 50 (1): 203-241. ULJÉ, C.B. & BAS, C., 1991. Studies in Coprinus II. Subsection Setulosi of section Pseudocoprinus. Persoonia 14(3): 275–339. ULJÉ, K., 2001. Coprinus site. [Data di accesso: 01/04/2021]. WÄCHTER, D. & MELZER, A., 2020. Proposal for a subdivision of the family Psathyrellaceae based on a taxon-rich phylogenetic analysis with iterative multigene guide tree. Mycological Progress 19: 1151–1265. https://doi.org/10.1007/s11557-020-01606-3. Scheda realizzata da Sergio Mombrini e Tomaso Lezzi. Microscopia di Segio Mombrini. Regione Lombardia; Marzo 2021; Foto macro- e micro- scopiche di Sergio Mombrini. Ritrovamento su letamaio. Esemplari in habitat. Spore. Osservazione in Rosso Congo ammoniacale, a 1000×. Basidi. Osservazione in Rosso Congo ammoniacale, a 400×. Cheilocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Pleurocistidio. Osservazione in Rosso Congo ammoniacale, a 400×. Pileipellis e pileocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Pileocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Caulocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Ife miceliari. Osservazione in Rosso Congo ammoniacale, a 400×. -
Coprinopsis cordispora (T. Gibbs) Watling & M.J. Richardson 2020
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
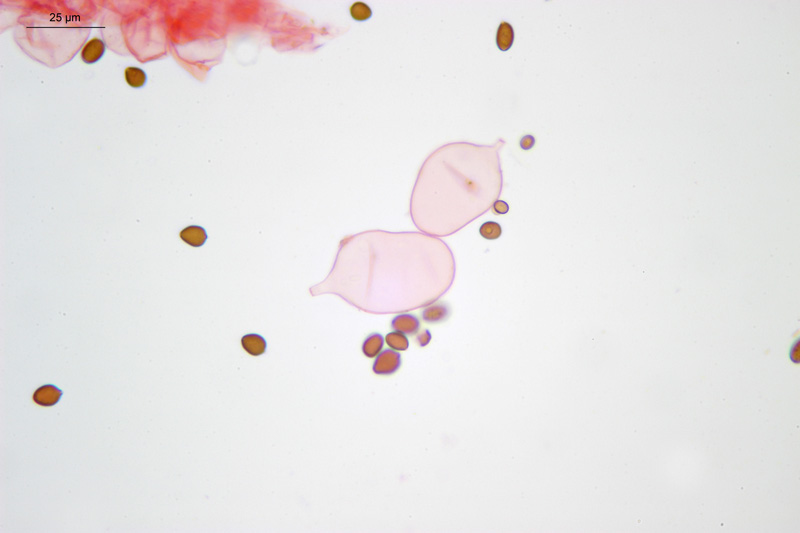
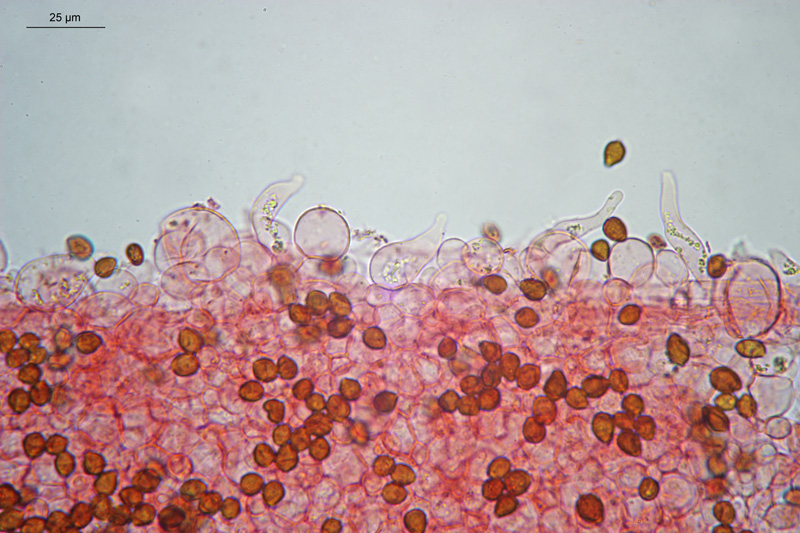
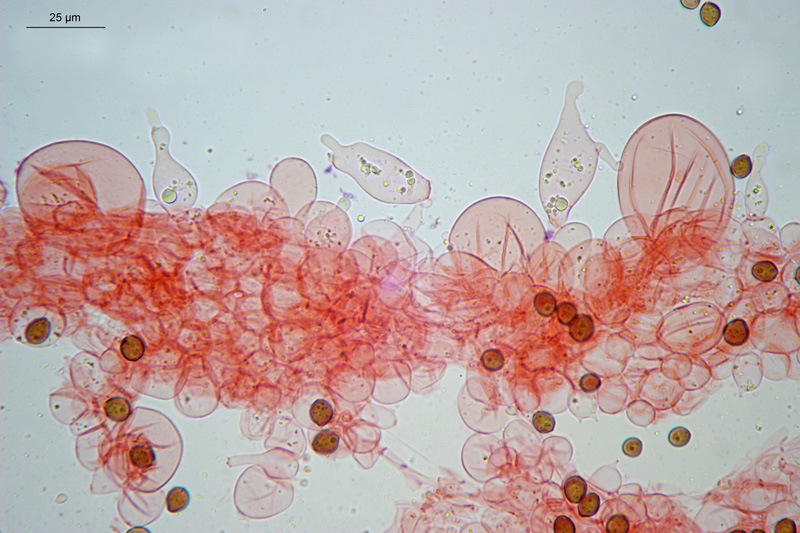
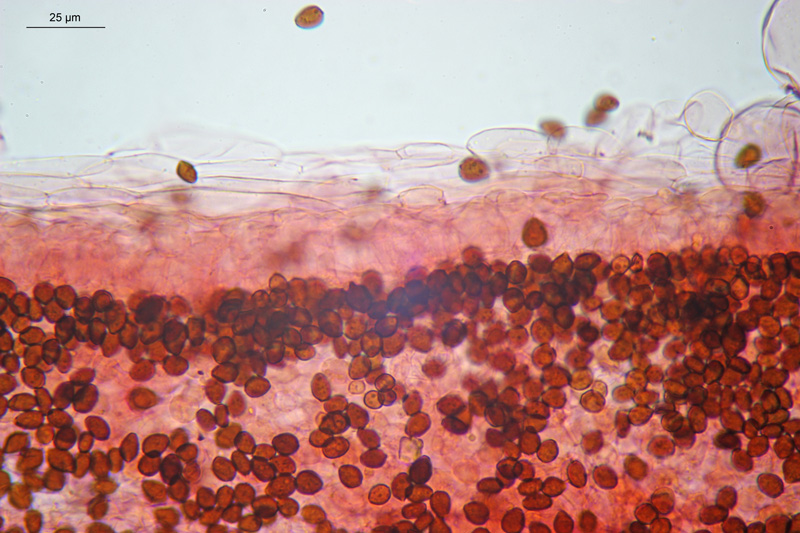
Coprinopsis cordispora (T. Gibbs) Watling & M.J. Richardson 2020 Tassonomia Divisione Basidiomycota Classe Agaricomycetes Ordine Agaricales Famiglia Psathyrellaceae Sinonimi Coprinopsis patouillardii Quél. s. auct. p.p. 1883 Coprinus volvaceominimus Crossl. [come 'volvaceo-minimus'] 1891 Coprinus cordisporus T. Gibbs, Naturalist 1908 Narcissea cordispora (T. Gibbs) D. Wächt. & A. Melzer 2020 Etimologia L'epiteto Coprinopsis deriva dal genere Coprinus con il suffisso -ópsis dal greco ὄψις [ópsis] = dall'aspetto di, per l'aspetto simile a Coprinus. L'epiteto cordispora deriva dal latino cors-cordis = cuore, e spora, per le spore a forma di cuore. Cappello Chiuso con forma ellissoidale-cilindrica, aperto con diametro di circa 15-20 mm, coperto da un velo fioccoso, bianco. Lamelle Libere al gambo, presto grigie e infine nere e deliquescenti. Gambo 30-40 × 1 mm, ialino, liscio, sono visibili solo alcune fibrille, alla base presenta un piccolo bulbillo. Carne Esigua, odore assente. Habitat Fungo coprofilo. Microscopia Spore 9,3-11 × 7-8,7 µm; Qm= 1,31; lentiformi, più o meno cuoriformi in vista frontale, ellissoidali in vista laterale; poro germinativo centrale. Basidi 18-26 × 8-10 µm, tetrasporici. Pleurocistidi 35-58 × 18-30 µm, utriformi. Cheilocistidi 23-51 × 15-30 µm, subglobosi, ellissoidali, presenti anche lageniformi 25-43 × 7-15 µm. Epicutis costituita da ife allungate, ellissoidali, più o meno parallele. Velo costituito da sferociti, con parete un poco ispessita, lisci, ialini, di diametro 15-60 µm. GAF assenti. Pileocistidi (setae-setole) assenti. Caulocistidi assenti. Commestibilità e tossicità Non commestibile. Osservazioni Per quanto concerne la nomenclatura e alcune delle sinonimie si è fatto riferimento a Funga Nordica (2012), Index Fungorum utilizza anch'esso il binomio Coprinopsis cordispora, mentre Mycobank tenendo conto del recente lavoro di Dieter Wächter & Andreas Melzer (2020) aggiorna la nomenclatura a Narcissea cordispora. Specie simili Coprinopsis patouillardii (Quél.) Gminder, avrebbe spore simili nella forma ma più piccole: 6-9 × 6-8 µm, cheilocistidi esclusivamente subglobosi, crescerebbe in genere su cumuli di compost, paglia, vinaccia o scarti di verdure, alcuni autori adottando un concetto piuttosto ampio di questo taxon e vi comprenderebbero anche Coprinopsis cordispora. Secondo Uljé (2001) Il carattere più distintivo di Coprinopsis cordispora è la presenza di cheilocistidi lageniformi insieme a quelli subglobosi. Bibliografia AA.VV., 2012. Funga Nordica. Agaricoid, boletoid and cyphelloid genera. Ed. Nordsvamp. CACIALLI, G., CAROTI, V. & DOVERI, F., 1999. Contributio ad Cognitionem Coprinorum. Monografie di Pagine di Micologia 1: 1–256. CACIALLI, G., CAROTI, V. & DOVERI, F., 2002. Aggiornamento della Monografia "Contributio ad Cognitionem Coprinorum". Mycologia Montenegrina V, Separatum: 119–129. CITÉRIN, M. 1992. Clé analytique du genre Coprinus Pers. Doc. Mycol. 22(86): 1–28. ULJÉ, C.B. & BAS, C., 1991. Studies in Coprinus II. Subsection Setulosi of section Pseudocoprinus. Persoonia 14(3): 275–339. ULJÉ, K., 2001. Coprinus site. [Data di accesso: 01/04/2021]. WÄCHTER, D. & MELZER, A., 2020. Proposal for a subdivision of the family Psathyrellaceae based on a taxon-rich phylogenetic analysis with iterative multigene guide tree. Mycological Progress 19: 1151–1265. https://doi.org/10.1007/s11557-020-01606-3. Scheda realizzata da Sergio Mombrini e Tomaso Lezzi. Microscopia di Segio Mombrini. Regione Lombardia; Marzo 2021; Foto macro- e micro- scopiche di Sergio Mombrini. Ritrovamento su letamaio. Esemplari in habitat. Spore. Osservazione in Rosso Congo ammoniacale, a 1000×. Basidi. Osservazione in Rosso Congo ammoniacale, a 400×. Cheilocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Cheilocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Pleurocistidi. Osservazione in Rosso Congo ammoniacale, a 400×. Epicutis. Osservazione in Rosso Congo ammoniacale, a 400×. Velo. Osservazione in Rosso Congo ammoniacale, a 400×.


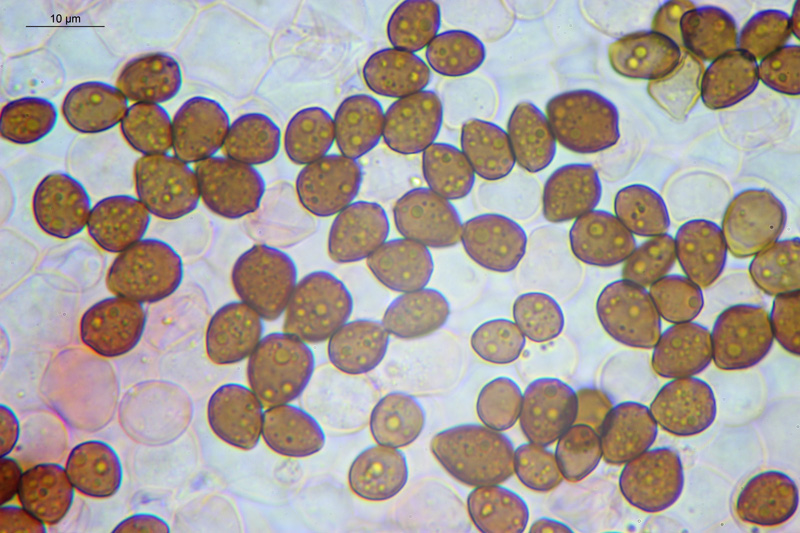
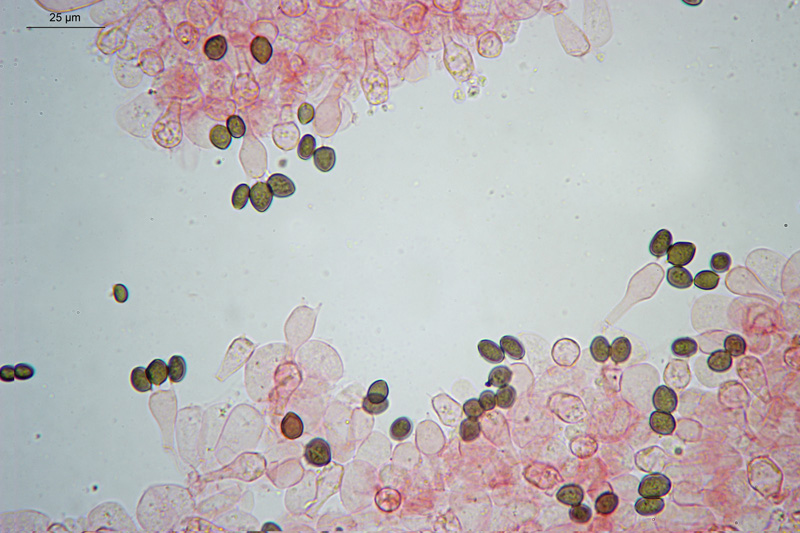
-
Trichoglossum hirsutum (Pers. : Fr.) Boud. 1907
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
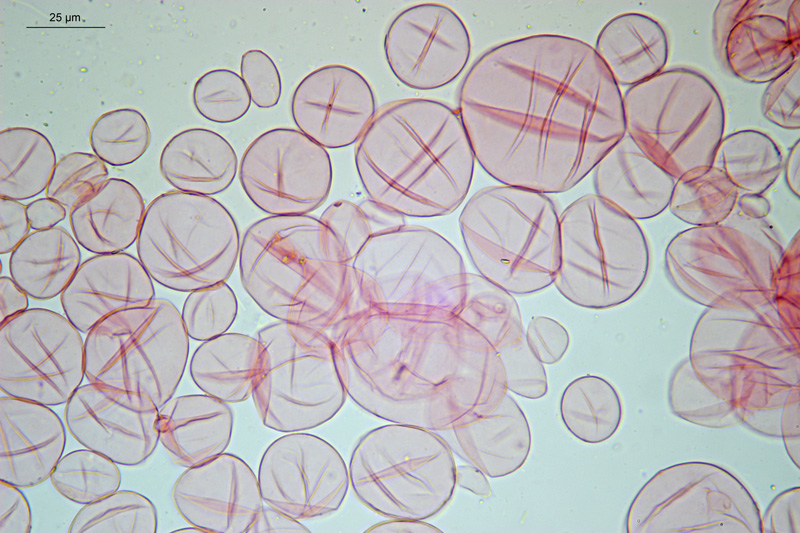
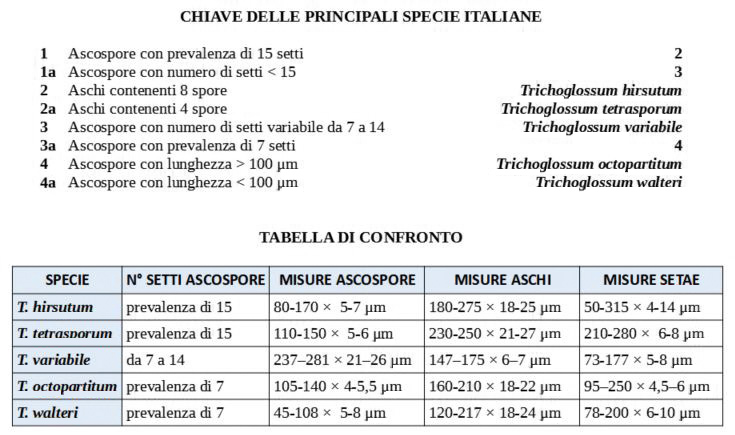
Trichoglossum hirsutum (Pers. : Fr.) Boud. 1907 Tassonomia Divisione Ascomicota Classe Leotiomycetes Ordine Helotiales Famiglia Geoglossaceae Sinonimi Trichoglossum hirsutum var. wrightii E. J. Durand 1908 Trichoglossum hirsutum var. doassansii Pat. 1909 Trichoglossum hirsutum var. capitatum (Pers.) Teng 1934 Trichoglossum hirsutum var. variabile (E. J. Durand) S. Imai 1941 Trichoglossum hirsutum var. heterosporum Mains 1954 Trichoglossum hirsutum var. irregulare Mains 1954 Trichoglossum hirsutum var. longisporum (F. L. Thay) Mains 1954 Trichoglossum hirsutum var. multiseptatum Mains 1954 Trichoglossum hirsutum var. latisporum W. Y. Zhuang 1997 Trichoglossum hirsutum var. caespitosum Lécuru 2013 Etimologia L'epiteto Trichoglossum deriva dal greco “trichós” = peli e da “glóssa” = lingua, per l'ascoma finemente villoso, linguiforme. Il termine hirsutum deriva da “hirsútus” = irsuto, ispido, in riferimento ai peli. Ascoma Apotecio con altezza complessiva fino a 60-70 mm, di color nero, con superficie caratteristicamente vellutata a causa dei piccoli peli che lo ricoprono totalmente, visibili agevolmente con una lente. Di forma slanciata e variabile, è composto da una clavula, che comprende la parte fertile, e uno pseudostipite, separati da una vallecola non sempre ben evidente. La clavula si presenta linguiforme, spatulata, molto spesso "a forma di picche", comunque sempre appiattita e solcata. Lo pseudostipite invece è per lo più cilindrico ed è lungo più del doppio rispetto alla clavula. Carne Compatta, di consistenza elastica, di color bruno-nerastro. Habitat Ambiente mediterraneo, umido, muscicolo, dall'autunno all'inverno inoltrato. Microscopia Ascospore (108,7) 112,7-131,8 (139,6) × (4) 4,6-5,7 (6,2) µm; Q = (19,1) 20,3-26,5 (33); N = 56; Media = 122 × 5,2 µm; Qm = 23,9; di forma cilindrica con le estremità attenuate, bruno-nerastre, fuliginose, con pareti e setti più scuri; pluriguttulate, con presenza di setti trasversali, in numero di 15 con rare eccezioni. Aschi (143,6) 166,9-210,6 (211,0) × (13,4) 13,41-17,58 (17,6) µm; Q = (10,3) 10,4-13,9 (15,7); N = 22; Media = 190,0 × 16,0 µm; Qm = 11,9; ottasporici con spore allungate longitudinalmente; sono inopercolati con poro amiloide e base pleurorinca. Parafisi ialine, cilindriche, filiformi, settate, spesso diverticolate; gli articoli terminali hanno forme notevolmente differenziate: cilindriche, subclavate-clavate, leggermente ricurve, a gancio, arrotolate, a martello, oppure con curvature irregolari, spesso presentano diverticoli anche molto pronunciati. Excipulum ectale formato da elementi catenulati, di varie forme e dimensioni. Setae (peli) di varie dimensioni, fino a 320 µm di lunghezza, di forma lanceolata, con apice acuto, di color bruno-nero, a parete spessa. Si trovano sia nella parte fertile (clavula) che nello pseudostipite, dove però risultano di lunghezza per lo più minore rispetto a quelle della clavula. Commestibilità e tossicità Non commestibile. Specie simili Il Genere Trichoglossum appartiene alla Famiglia delle Geoglossaceae che comprende specie terrestri, claviformi o capitate e stipitate, di colore scuro, spesso nerastro, pertanto macroscopicamente risultano difficilmente distinguibili tra loro. Per la loro forma particolare sono conosciute con il nome volgare di “Lingue di terra". I Generi Hemileucoglossum e Leucoglossum si differenziano per avere spore ialine, mentre i Generi Geoglossum e Glutinoglossum per la completa assenza di setae. Dopo aver approfondito il Genere Trichoglossum, proponiamo una nostra chiave dicotomica delle principali specie italiane ed una tabella di confronto che evidenzia le caratteristiche microscopiche peculiari di ciascuna specie Bibliografia ARAUZO, S., & IGLESIAS, P., 2014. La familia Geoglossaceae ss. str. en la península ibérica y la Macaronesia. Errotari 11: 166-259. BREITENBACH, J. & KRÄNZLIN, F., 1984. Fungi of Switzerland. Vol 1 Ascomycetes. Verrlag Mykologia, Lucerne. HLADKI, A.I & ROMERO, A.I., 2009. La familia Geoglossaceae s. str. (Helotiales) en la provincia de Tucumán (Argentina). Boletín de la Sociedad Argentina de Botánica. 44 (3-4): 249-255. KUČERA, V., LIZOŇ, P. & KAUTMANOVÁ, I., 2010. Geoglossoid fungi in Slovakia II. Trichoglossum octopartitum, a new species for the country. Czech Mycology. 62 (1): 13-18. MEDARDI, G., 2006. Atlante fotografico degli Ascomiceti d'Italia. Trento: Ed. AMB. PRIOU, J.P. & DECHAUME, J.P., 2014. Geoglossaceae de France: Trichoglossum octopartitum Mains, récolté en France. Ascomycete.org, 6 (5): 138-142. Scheda di proprietà AMINT realizzata da Laura Nicoletti e Manuel Atzeni. Approvata e revisionata dal CLR Micologico di AMINT. Regione Lazio; Gennaio 2021; Foto di Laura Nicoletti e Manuel Atzeni. I nostri esemplari sono stati trovati in habitat mediterraneo, in ambiente umido, muscicolo, dall'autunno all'inverno inoltrato. Aschi e parafisi. Osservazione in Reattivo di Melzer, a 400×. Aschi. Osservazione in Reattivo di Melzer, a 400×. Aschi e setae. Osservazione in Reattivo di Melzer, a 400×. Asco. Osservazione in Reattivo di Melzer, a 400×. Aschi. Osservazione in Reattivo di Melzer, a 400×. Parafisi. Osservazione in acqua, a 600×. Spore. Osservazione in acqua, a 600×. Setae pseudostipite. Osservazione in Rosso Congo ammoniacale, a 100×. Excipulum ectale. Osservazione in Rosso Congo ammoniacale, a 200×. Excipulum ectale. Osservazione in Rosso Congo ammoniacale, a 200×.
-
Peziza saccardoana Cooke 1877
Archivio Micologico ha risposto alla discussione di Archivio Micologico in Funghi Non Commestibili o Sospetti
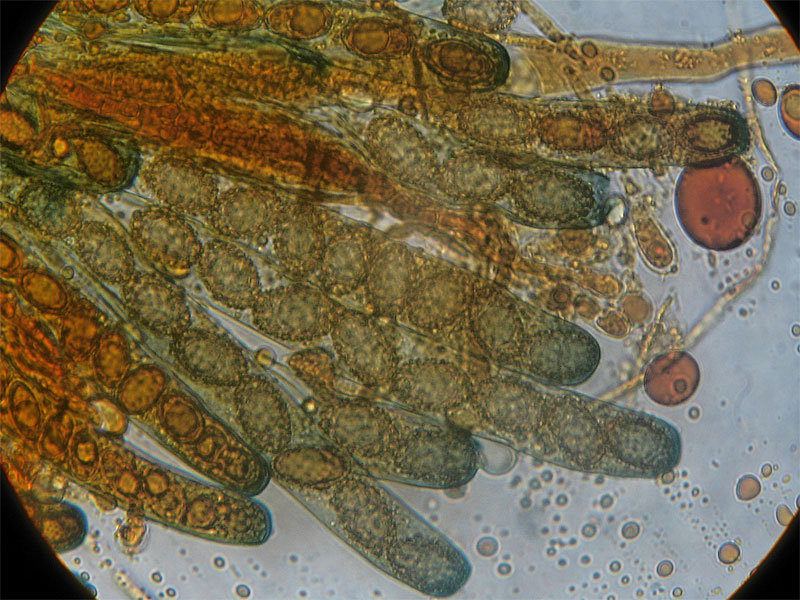
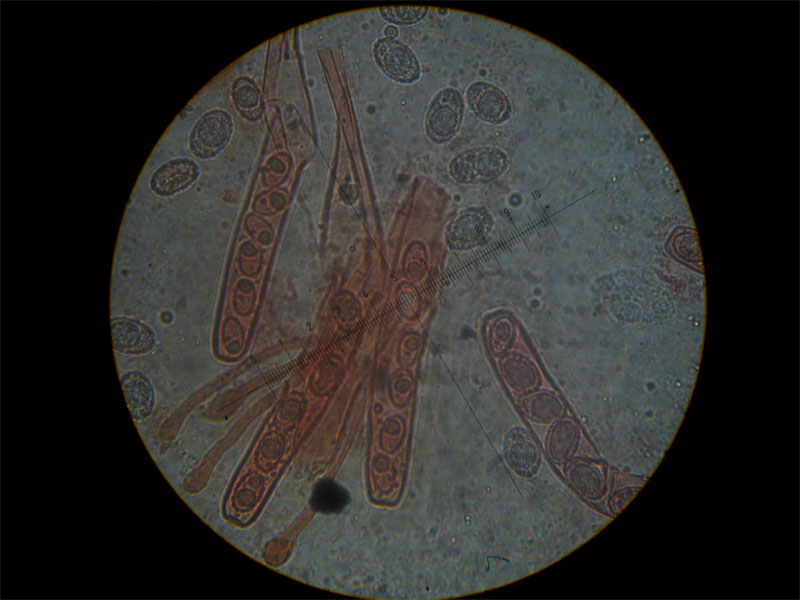
Peziza saccardoana Cooke 1877; Regione Sardegna; Gennaio 2008; Foto di Giovanni Satta. In un rado querceto di pianura, con sottobosco di Cistus monspeliensis e Lentisco. Assenza di latice alla frattura, crescita con esemplari appressati, superficie esterna particolarmente rugolosa. Regione Sardegna; Gennaio 2008; Foto di Franco Sotgiu. Studio microscopico e foto di Massimo Biraghi. Aschi ottasporici uniseriati , parafisi cilindracee con apice allargato a volte fino a 5-7 µm. Spore ellissoidali, monoguttulate, verrucose mediamente di 15 × 10 µm. Osservazione in melzer. Aschi e spore. Aschi che arrivano fino a 380 µm. Spore ellissoidali e verrucose. Blu cotone in lactofenolo a freddo.



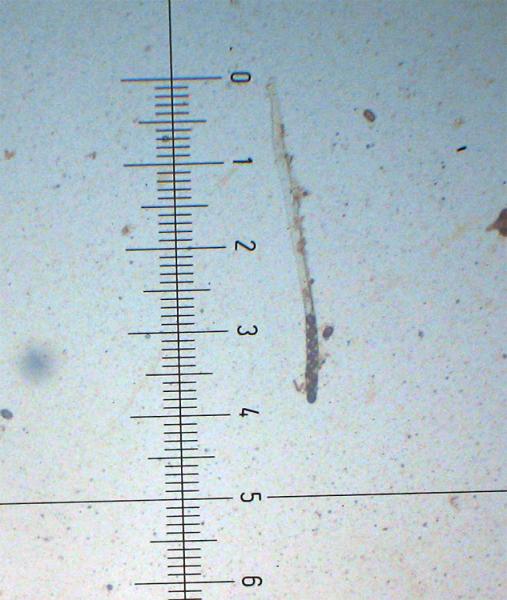
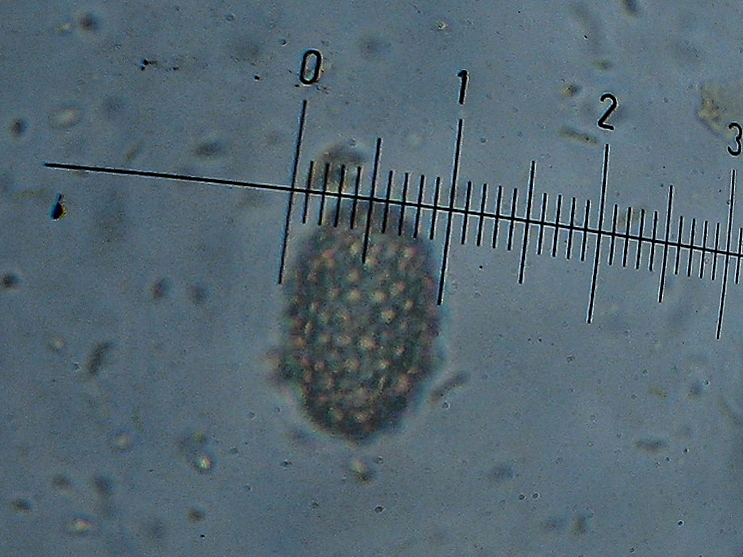
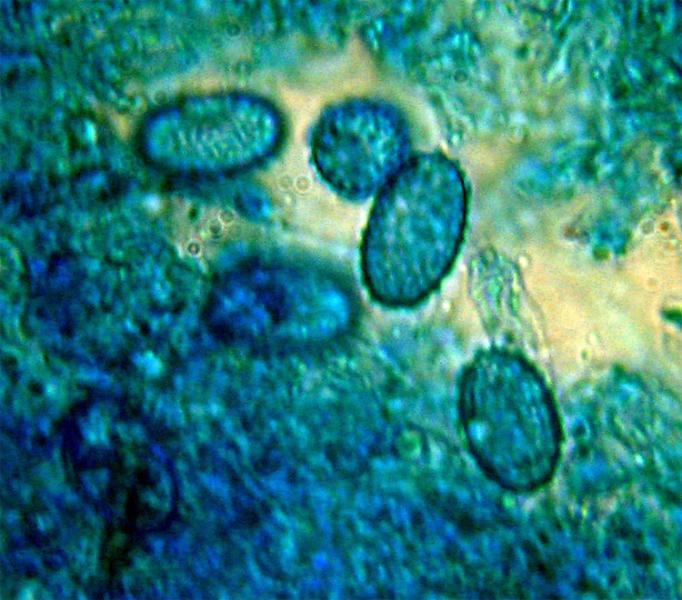
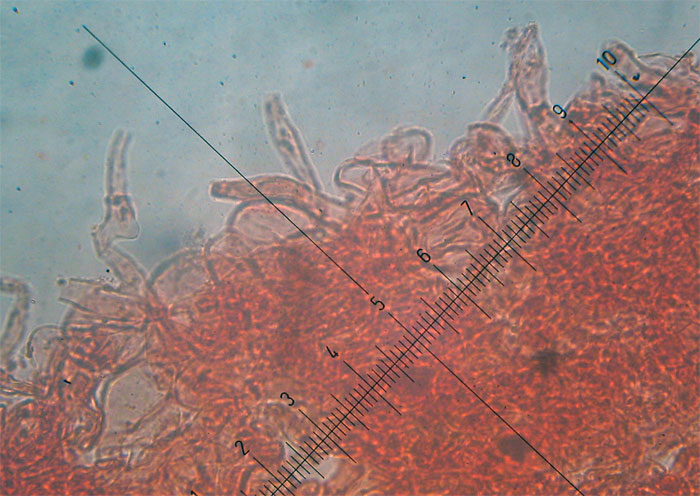
-
Pseudoplectania ericae Donadini, 1987
Archivio Micologico ha postato un topic nell'area Funghi Non Commestibili o Sospetti
Pseudoplectania ericae Donadini 1987 Tassonomia Divisione Ascomycota Classe Pezizomycetes Ordine Pezizales Famiglia Sarcosomataceae Sinonimi Plectania ericae (Donadini) Roqué 2009 Etimologia L'epiteto Pseudoplectania = "falsa Plectania" deriva dal sostantivo greco “pseudo” = falso, menzognero, ingannatore, simile all’apparenza, fallace, e da Plectania, dal greco "πλεκτάνη" [plektàne] = intreccio, attorcigliamento, groviglio. L'epiteto ericae è riferito allo stretto legame con Erica spp.. Ascoma Apotecio con diametro 2-10 mm, sessile, inizialmente cupuliforme tendente a distendersi a maturità. Superficie interna imeniale liscia, di color nero lucido, superficie esterna concolore ma opaca, tomentosa, ricoperta da una fine peluria. Carne Nerastra, di consistenza gelatinosa. Habitat In areale mediterraneo, in associazione con Erica spp.. Microscopia Spore (11,2) 11,4-12,8 (13) × (10,8) 11,2-12,5 (12,7) µm; Q = 1 (1,1); N = 32; Media = 11,9 × 11,7 µm; Qm = 1; a parete spessa, sferiche, lisce, con piccole guttule all'interno. Si nota che spesso il diametro delle spore all'interno degli aschi è maggiore del diametro di questi. Aschi (266,8) 303,6-333 (340,2) × (10,1) 11,1-14.06 (14,1) µm; Q = 22,2-29,2 (32,9); N = 10; Media = 313,5 × 12,1 µm; Qm = 26,2; cilindrici, inamiloidi, ottasporici. Parafisi cilindriche, filiformi, settate, biforcate all'apice, con un contenuto granuloso giallo scuro, specialmente dalla metà superiore fino all'apice. Excipulum medullare formato da una textura intricata di ife cilindriche. Excipulum ectale formato da una textura angularis, con elementi cilindrici/globulosi; si notano sia peli corti a parete spessa, che peli molto più lunghi con parete meno ispessita, entrambi settati, da ondulati a lisci. Commestibilità e Tossicità Non commestibile. Osservazioni Possiamo differenziare il Genere Plectania dal Genere Pseudoplectania per la forma sporale: in Plectania troviamo spore ellissoidali, mentre in Pseudoplectania le spore sono sferiche. Specie simili Pseudoplectania nigrella (Pers. : Fr.) Fuckel; cresce in primavera, inizio estate, in zone montane con conifere, principalmente Picea spp.. È sessile come Pseudoplectania ericae, ma gli ascomi sono più grandi, possiede parafisi con apice leggermente dilatato, spesso lobato, diverticolato e i peli dell’excipulum ectale sono tortuosi, flessuosi, molto intrecciati, a differenza di quelli di Pseudoplectania ericae che sono lisci e non arrotolati. Pseudoplectania melaena (Fr. : Fr.) Sacc.; è legata principalmente ad Abies spp.. A differenza di Pseudoplectania ericae è cortamente stipitata, le parafisi sono più o meno diverticolate, nodulose e ricurve, uncinate all’apice. Pseudoplectania episphagnum (J. Favre) M. Carbone, C. Agnello & P. Alvarado; specie rara, anch’essa di piccole dimensioni come la Pseudoplectania ericae ma ha un’ecologia completamente diversa, infatti è associata a Sphagnum spp.. Sebbene abbia una microscopia molto simile a Pseudoplectania nigrella si differenzia da questa, oltre che per l’habitat, anche per possedere un corto stipite. Bibliografia BOCCARDO F., CARBONE M., & VIZZINI A., 2014. Pseudoplectania ericae, una rara specie rinvenuta in Liguria (Italia). CARBONE, M., AGNELLO, C. & ALVARADO P., 2014. Phylogenetic and morphological studies in the genus Pseudoplectania (Ascomycota,Pezizales). CARBONE, M., AGNELLO, C. & BAGLIVO A., 2010. Appunti su Plectania rhytidia e studio del typus di Urnula platensis. Rivista di Micologia, 53 (2): 119-135. CARBONE M. & AGNELLO C., 2012. Studio e tipificazione di Pseudoplectania nigrella. MEDARDI, G., 2006. Atlante fotografico degli Ascomiceti d'Italia. Trento: Ed. AMB. PÉREZ-DE-GREGORIO, M. A., 2009. Algunos hongos interesantes de Girona. Fungi non Delineati. 44: 43. VAN VOOREN, N., 2014. Contribution à la connaissance des Pézizales (Ascomycota) de Rhône-Alpes 2° partie. Scheda di proprietà AMINT realizzata da Laura Nicoletti e Manuel Atzeni. Approvata e revisionata dal CLR Micologico di AMINT. Regione Toscana; Gennaio 2020; Foto di Laura Nicoletti e Manuel Atzeni. (Exsiccata MA-486 ; LN_20200115_12) Sezione. Osservazione in acqua, a 40×. Aschi e parafisi. Osservazione in Blu cotone in acido lattico, a 400×. Aschi, spore, parafisi. Osservazione in Blu cotone in acido lattico, a 400×. Aschi, spore, parafisi. Osservazione in Rosso Congo ammoniacale, a 400×. Aschi, spore, parafisi. Osservazione in Melzer, a 400×. Aschi, spore, parafisi. Osservazione in acqua, a 1000×. Aschi, spore, parafisi. Osservazione in Blu di Toluidina, a 1000×. Aschi, spore, parafisi. Osservazione in campo oscuro, a 400×. Peli excipulum ectale. Osservazione in Blu cotone in acido lattico, a 100×. Peli excipulum ectale. Osservazione in acqua, a 400×.